© 2002 Nature Publishing Group http://neurosci.nature.com
brief communications
Deficits in spatial coding and feature binding following damage to spatiotopic maps in the human pulvinar Robert Ward1, Shai Danziger1,2, Vanessa Owen1 and Robert Rafal1 1 Centre for Cognitive Neuroscience, University of Wales, Bangor LL57 2AS, UK 2 Ben Gurion University, PO Box 653, Beer Sheva, Israel
Correspondence should be addressed to R.W. (
[email protected])
Published online: 7 January 2002, DOI: 10.1038/nn794
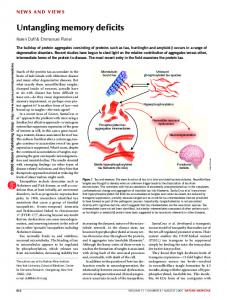
We report a patient with unilateral damage to the rostral part of the pulvinar who was impaired in localizing stimuli in the inferior visual field contralateral to the lesion and who made errors in the binding of shape and color in that quadrant. The findings demonstrate the importance of the pulvinar in spatial coding and provide support for the function of the thalamus in binding of features1. They also provide evidence for a homology between the visual field maps of the inferior and lateral subdivisions of the pulvinar in monkeys and in humans, such that the inferior visual field is represented in the rostral part of the nucleus.
Visual processing is distributed across specialized brain regions, yet visual experience is integrated and cohesive. Coordinating multiple visual processors to represent a single object has been described as the ‘binding problem’2. We studied the breakdown of feature binding in patient T.N. by measuring illusory conjunctions, in which visual features are correctly perceived but incorrectly combined. An observer might, for example, report seeing a blue X when shown a blue O and a pink X3. Accurate binding depends on accurate spatial coding, so features from nearby objects are more likely to be conjoined than those from distant ones4, and binding in the periphery is especially difficult5. T.N. suffered a unilateral hemorrhage in the posterior thalamus extending into the rostral part of the pulvinar 8 months before testing (Fig. 1). The damaged region corresponds to the location of the inferior and lateral maps found in the monkey pulvinar. In the monkey, these maps exhibit a clear spatiotopic organization, with the lower visual field represented dorsally, and the upper field, ventrally6–8. We wished to see whether damage to this area in humans would produce deficits in spatial coding and/or the use of spatial information in binding. Based on monkey neuroanatomy, we expected T.N.’s lesion at the dorsal and anterior boundary of the right pulvinar to produce deficits specific to the lower left visual quadrant. T.N. was asked to report the identity and color of a target letter (either an F or an X) while ignoring a distractor letter (an O) appearing within the same hemifield (Fig. 2). Target and distractor appeared in four possible colors, although never both in the same color during one trial. Our primary interest was the target color report. Two types of errors were possible. First, the target could be misidentified with the color of the distractor. These
a
b
Fig. 1. Reconstructed T1-weighted magnetic resonance images from patient T.N., showing a lesion involving the posterior-lateral thalamus and the most anterior and dorsal part of the pulvinar. The right hemisphere is on the left side in all pictures. (a) Axial slices. Intact pulvinar enclosed by white boxes. The anterior border of the pulvinar is approximated by the plane perpendicular to the posterior commissure (PC)11; in slices from 0 to +8 mm, the lesion encroaches on this landmark. (b) Coronal slices. Involvement of the dorsal pulvinar is seen in slices from –2 to 0 mm from posterior commissure perpendicular; the intact pulvinar curves convexly into the ventricle, but the damaged pulvinar does not (white arrows). T.N.’s sensation and mobility in the left side and limbs were moderately impaired. Extraocular movements were normal, visual fields were intact, and there was no visual or tactile extinction on confrontation. nature neuroscience • volume 5 no 2 • february 2002
99
© 2002 Nature Publishing Group http://neurosci.nature.com
brief communications
Fig. 2. Sample stimulus display. The target was either an F or an X; the distractor was always an O. Letters appeared in one hemifield symmetrically about the horizontal meridian, and were colored red, green, blue or yellow (a black background was used in testing), but were always different colors. The two white digits, one small and one large, were presented centrally. Digits were selected at random from 1 to 4 but were never the same. Identifying the physically larger digit was the primary task; this has previously been shown to increase the rate of illusory conjunctions3. Stimulus displays appeared briefly and were masked (mean exposures, experiment 1, 140 ms; experiment 2, 108 ms; experiment 3, 89 ms). Viewing distance was approximately 50 cm but not fixed. In experiments 1 and 2, T.N. was asked to report, in order, the larger digit, the target color and the target identity. In experiment 3, T.N. additionally reported the location (up or down) of the target. Responses were unspeeded key presses. Accuracy on the primary task was 98%; target identification (F or X) was accurate in both contralesional (95%) and ipsilesional fields (99%).
conjunction errors demonstrate an illusory conjunction of form (from the target) and color (from the distractor). Second, one of the two colors not appearing in the display could be reported as the target color. These feature errors indicate difficulty with color perception rather than binding. In experiment 1, we observed more conjunction errors in contralesional than ipsilesional fields (contralesional, 19%, 48/255 trials; ipsilesional, 2%, 4/251; χ21 = 40.7, p < 0.00005). Crucially, this increase was not the result of difficulty in color and lateral maps of the monkey. We conclude that damage to discrimination, as feature error rates were low and equivalent spatiotopic maps within the pulvinar degrades the spatial codbetween fields (contralesional, 2%, 5/255; ipsilesional, 3%, ing of visual features. In the presence of nearby distractors, fea7/251). In experiment 2, performance on ‘near’ displays in which tures in T.N.’s impaired quadrant were more likely to be target and distractor were relatively close together (1.7°) was mislocalized and so more likely to be incorrectly bound to feacompared with that on far displays in which they were further tures at nearby locations. Spatiotopic maps in the pulvinar, along apart (5.1°). Contralesional conjunction errors were reduced in with parietal cortex9,10, seem to be part of a distributed network far compared with near displays (far, 13%, 14/105; near, 32%, determining the location of features and their conjunctions. 35/110; χ21 = 10.4, p = 0.001). If anything, contralesional feature errors showed the reverse trend, being nonsignificantly Acknowledgements higher in the far condition (6%, 6/105 trials) than in the near This work was supported by a grant from the Medical Research Council (UK). (2%, 2/110) (χ21 = 2.28, p = 0.13). Finally, across both experiThe research was approved by the Ethics Comittees of the University of Wales, ments, conjunction errors were more frequent in the lower conBangor, the North West Wales NHS Trust and the Conwy–Denbighshire NHS tralesional quadrant than any other (χ21 > 30.0, p < 0.00005) Trust. We thank T.N. for her help in these studies. with no difference in feature errors between quadrants RECEIVED 19 AUGUST; ACCEPTED 7 DECEMBER 2001 (χ21 < 1.5, p > 0.2). These results suggest a deficit in forming discriminable spatial codes for features from nearby objects, particularly for targets in the lower contralesional quadrant. 1. Crick, F. Proc. Natl. Acad. Sci. USA 81, 4586–4590 (1984). We reasoned that if T.N.’s contralesional conjunction errors were 2. Treisman, A. Curr. Opin. Neurobiol. 6, 171–178 (1996). 3. Treisman, A. & Schmidt, H. Cogn. Psychol. 14, 107–141 (1982). caused by a degraded spatial coding of features, her ability to local4. Cohen, A. & Ivry, R. J. Exp. Psychol. Hum. Percept. Perform. 15, 650–663 ize targets might also be impaired. Experiment 3 was identical to (1989). 5. Prinzmetal, W., Henderson, D. & Ivry, R. J. Exp. Psychol. Hum. Percept. experiment 1 except that T.N. also reported the upper or lower locaPerform. 21, 1362–1375 (1995). tion of the target. Mislocalizations were most frequent for targets 6. Bender, D. B. J. Neurophysiol. 46, 672–693 (1981). in the lower contralesional quadrant, occurring on 25% (51/203) 7. Petersen, S. E., Robinson, D. L. & Keys, W. J. Neurophysiol. 54, 867–886 (1985). of trials, compared with a rate of 4% (8/206) or less in the other 8. Robinson, D. L. & McClurkin, J. W. in The Neurobiology of Saccadic Eye three quadrants (χ21 > 37.0, p < 0.000005). Pairwise comparisons Movements (eds. Wurtz, R. H. & Goldberg, M. E.) 337–360 (Elsevier, New between other quadrants did not suggest any reliable difference in York, 1989). 2 localization (χ 1 < 1.4, p > 0.2). Feature and conjunction error rates 9. Cohen, A. & Rafal, R. D. Psychol. Sci. 2, 106–110 (1991). 10. Friedman-Hill, S. R., Robertson, L. C. & Treisman, A. Science 269, 853–855 were conditionalized on target localization accuracy (Table 1). Con(1995). junction errors were more likely when the target was mislocalized 11. Talairach, J. & Tournoux, P. Co-Planar Stereotaxic Atlas of the Human Brain than when it was correctly localized (79% versus 4%, respectively; (Thieme, New York, 1988). χ21 = 385.8, p < 0.000005). Conjunction error rates for mislocalized Table 1. Results from experiment 3, showing target mislocalizations, feature and targets were relatively constant conjunction errors, conditionalized on location accuracy. across quadrants; what varied Number of trials Number of feature Number of conjunction between quadrants was the likelierrors and rate (%) errors and rate (%) hood of a mislocalization in the first Contralesional Ipsilesional Contralesional Ipsilesional Contralesional Ipsilesional place. T.N.’s impairment in feature binding therefore seems linked to a Correct deficit in target localization. Upper 198 203 8 (4) 12 (6) 16 (8) 4 (2) These results are the first eviLower 152 199 3 (2) 12 (6) 8 (5) 2 (1) dence for spatiotopic maps at the Incorrect rostral boundary of the human Upper 8 4 1 (13) 0 (0) 4 (50) 3 (75) pulvinar, homologous in location Lower 51 5 2 (4) 0 (0) 43 (85) 4 (80) and organization to the inferior 100
nature neuroscience • volume 5 no 2 • february 2002