Int J Primatol DOI 10.1007/s10764-015-9882-9
Defining Genera of New World Monkeys: The Need for a Critical View in a Necessarily Arbitrary Task Guilherme Siniciato Terra Garbino 1,2,3
Received: 12 August 2015 / Accepted: 19 October 2015 # Springer Science+Business Media New York 2015
Abstract The genus is one of the most important taxonomic categories because it appears in the two-part species name, is easier to identify than the species, and is very commonly the least inclusive category employed in paleontological, ecomorphological, phylogenetic, evolutionary, and biogeographic comparisons. However, the theoretical and practical definitions of genera have received little attention in the New World monkey literature. Six genus-level changes in platyrrhine taxonomy have been suggested since 1998. The genera Leontocebus, Mico, Oreonax, and Sapajus, have been revalidated and Callibella and BTorquatus^ proposed as new. Divergence time is becoming the favored criterion when defining genera, but most molecular phylogenetic studies date the divergence between alleles, not between species, which is the actual date of interest when studying systematics. I here review the recent genus-level changes in platyrrhine taxonomy and propose that we adopt a Bfocused monophyly^ approach for defining genera that overlays a wide variety of morphological, behavioral, and ecological characteristics on the molecular phylogeny to produce a classification. This proposal aims to facilitate communication among researchers and to offer a robust, more predictive, taxonomy that will better withstand future investigations. Keywords Broad monophyly . Divergence time . Focused monophyly . Genus concept . Platyrrhini . Taxonomy
* Guilherme Siniciato Terra Garbino
[email protected] 1
Pós-graduação em Zoologia, Universidade Federal de Minas Gerais, 31270-901 Belo Horizonte, Minas Gerais, Brazil
2
IPÊ - Instituto de Pesquisas Ecológicas, 12960-000 Nazaré Paulista, São Paulo, Brazil
3
Pós-graduação, Museu de Zoologia, Universidade de São Paulo, 04263-000 São Paulo, SP, Brazil
G. S. T. Garbino
Mere opinions, in fact, were as likely to govern people’s actions as hard evidence, and were subject to sudden reversals as hard evidence could never be. —Kurt Vonnegut, Galápagos
Introduction The genus is the first step in the hierarchical ladder of higher taxonomic levels, and has special properties that distinguish it from both the less inclusive species group and the more inclusive categories (Rosenberger 2012; Simpson 1961; Tattersall 2014). First, it differs from all more inclusive categories (family group names and above) by appearing in the scientific name of a species (Mayr 1943), which is formed by a specific epithet and a generic substantive (Dubois 2000). This makes the genus more relevant than the other higher categories when taxonomic stability is concerned. When used in a specific name, the genus name gives the idea of an exclusive group of related units, while the species name expresses the uniqueness of the unit (Cain 1958). Second, the genus is perhaps the easiest category to pinpoint without further investigation, in the field and among museum specimens (Anderson 1940; Atran 1987; Rosenberger 2012; Simpson 1943). This fact, together with the deeper time span of genera, when compared with species, has led some authors to consider it as one of the most operationally useful taxonomic categories (Allmon 1992; Simpson 1945; Tattersall 2014). Therefore, the genus is the most commonly used taxonomic category when comparing paleontological, ecomorphological, phylogenetic, evolutionary, and biogeographic data (Ayres and Clutton-Brock 1992; Ford and Davis 2009; Garber 1992; Hershkovitz 1977; Kay et al. 2008; Marroig and Cheverud 2005; Rosenberger 1992, 2011; Schneider and Sampaio 2015). Despite its relevance both in taxonomic practice and in communication, the genus is rarely discussed among zoologists in general (Dubois 1982, 1988). In primatology in particular, whereas many recent papers have discussed the species concept (Groves 2004, 2014a, b; Rosenberger 2014; Rylands and Mittermeier 2014; Tattersall 2007, 2014; Zinner and Roos 2014), the importance and singularity of the genus has been mentioned only briefly (Groves 2004; Rosenberger 2012; Tattersall 2014; Zinner and Roos 2014). Since 1998, two new genus group nomina have been proposed for platyrrhines: BTorquatus^ (Goodman et al. 1998) and Callibella (Van Roosmalen and Van Roosmalen 2003). Other authors have proposed the revalidation of four genera: Sapajus Kerr, 1792 (proposed as a subgenus by Silva-Junior 2001 and as a genus by Lynch-Alfaro et al. 2012a, b); Leontocebus Wagner 1839 (proposed by Buckner et al. 2015; Matauschek 2010; and Sampaio et al. 2015), Mico Lesson 1840 (first proposed as a subgenus by Goodman et al. 1998 and a genus by Rylands et al. 2000), and Oreonax Thomas, 1927 (proposed by Groves 2001). These names, however, were not widely accepted and the validity of some of them has been controversial (Aguiar and Lacher 2003, 2009; Garbino 2015; Rosenberger and Matthews 2008; Ruiz-García et al. 2014; Schneider et al. 2012). This disagreement derives, on the one hand, from the way in which the taxa were proposed, based on
Defining Genera of New World Monkeys
either low-support and poorly sampled phylogenies (Groves 2001; Van Roosmalen and Van Roosmalen 2003) or on divergence time alone (Buckner et al. 2015; Matauschek 2010), and, on the other hand, from differences in the view of what kind of information a generic name should communicate. Recent discussions have shifted the focus from questions such as BWhat should a genus be?^ or BWhat is the goal of a classification?^ to simply BHow do we rank the obtained phylogeny?^ (de Queiroz and Gauthier 1990; Dubois 2007; Goodman et al. 1998; Groves 2001, 2004; Sereno 1999). I here critically review the recent (post-1990) genus-level changes in New World monkey classification and recommend the revival of the classic character-based procedure for defining platyrrhine genera.
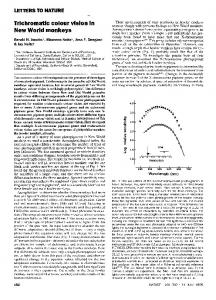
The Establishment and Treatment of Genera in New World Monkeys: The Concepts Behind the Practice The new genus-level proposals in platyrrhine taxonomy increased the number of generic names from the 16 traditionally recognized genera (Rosenberger 2012) to 22 genus-group taxa (21 genera plus a new subgenus). Three of the six nomenclatural changes proposed at the genus level refer to callitrichines (marmosets and tamarins). Although there is an obvious tendency for species number to increase in time among callitrichines, there is no clear relationship between the number of genera recognized and the number of species added to the group over the last 200 years (Fig. 1). The limited amount of material available for study, uncertainties about its provenance, and the difficulties in examining it personally suggest that, for most of the 19th century and the first half of the 20th century, necessarily arbitrary opinions were more influential than observable evidence in the classification of the marmosets and tamarins. Some authors chose to emphasize the many unique phenotypes found in the group, while other authors focused on the characters uniting the higher groups formed by the known species. Among the former, classifications with as many as seven genera were common (Gray 1870; Thomas 1922), while the latter authors subsumed all callitrichine diversity into, for example, a single genus with three subdivisions (Wagner 1839) or two genera (Geoffroy Saint-Hilaire 1812; Gray 1825). It was only after the first volume of Hershkovitz’s impressive Living New World Monkeys (1977) appeared that a robust multisource dataset was used to produce a classification of the Callitrichinae. This five-genera (or four if Cebuella is included in Callithrix) arrangement was historically the most stable one (Fig. 1), because it was supported by a collection of characters, several of which also represent synapomorphies of the four major, consensually recognized Callitrichidae lineages: marmosets (currently represented by Callithrix, Cebuella, and Mico), tamarins (Leontocebus and Saguinus), lion tamarins (Leontopithecus), and Goeldi’s monkey (Callimico) (Buckner et al. 2015; Garbino 2015; Hershkovitz 1977; Rosenberger 1984; Schneider and Sampaio 2015; Schneider et al. 1993). Reviewing the recent genus-level taxonomic proposals for New World monkeys reveals five common criteria for defining genera that are not mutually exclusive (Table I).
G. S. T. Garbino
60
50
40
30
20
10
0 1820
1840
1860
1880
1900
1920
1940
1960
1980
2000
2020
Year
Fig. 1 Number of species and subspecies (solid black line), genera (dash-dot line), and subgenera (dashed line) of callitrichines by year, based on Geoffroy Saint-Hilaire (1812), Wagner (1839), Lesson (1840), Reichenbach (1862), Gray (1870), Elliot (1913), Thomas (1922), Cabrera (1958), Hershkovitz (1977), Mittermeier et al. (1988), Rylands et al. (2000), Groves (2005), and Rylands and Mittermeier (2013).
Broad Monophyly-Based Criterion There is no doubt that Bgenera should be strictly monophyletic^ (Dubois 1982, p. 57). Expanded to all Linnaean taxonomic categories, this is the basis of synapomorphybased cladistic classification (Groves 2004; Hennig 1965; Platnick 1979; Wiley 1981), which clearly establishes monophyly as a mandatory criterion when defining genera or higher level taxa (Groves 2004, p. 1115). However, a classification that Bplace[s] species that are phenotypically very different in large genera solely on the basis of monophyly^ (Bbroad monophyly^ sensu Isler et al. 2013, p. 469) can be essentially arbitrary and uninformative. As an example, one could recognize as many as six different genera with few species or just a single genus with many species based on the clades in a recent phylogeny of the marmosets (Garbino 2015) (Fig. 2A). One of the first recent generic reshufflings of New World monkey taxonomy occurred because some studies nested Cebuella pygmaea within Callithrix, making this genus paraphyletic (Barroso et al. 1997; Porter et al. 1997; reviewed in Cortés-Ortiz 2009). This led Rylands et al. (2000) to restore Mico Lesson 1840 for the large-bodied Amazonian marmosets. To address the same situation, other authors classified all marmosets in a single genus, Callithrix, with Cebuella and Mico as subgenera (Goodman et al. 1998; Groves 2001, 2005), or lumped all marmosets in a single genus, Callithrix (Barroso et al. 1997).
Thomas (1927)
Kerr (1792)
Oreonax
Sapajus
X
Not based on systematic surveys (Van Roosmalen and Van Roosmalen 2003).
Nomen nudumd
Revalidated
X
Xa
X
X
X
X
X
X
Xc
X
Xb
X
X
Adaptive Divergence Morphological Broad Focused zone time gap monophyly monophyly
Underlying criteria
Goodman et al. (1998)
Lynch-Alfaro et al. (2012a, b); Silva-Júnior (2001)
Groves (2001)
Barroso et al. (1997); Garbino (2015); Porter et al. (1997); Rylands et al. (2000)
Buckner et al. (2015); Matauschek (2010); Sampaio et al. (2015)
Aguiar and Lacher (2003); Ford and Davis (2009); Van Roosmalen and Van Roosmalen (2003)
Source
d
Based on low-supported clades (