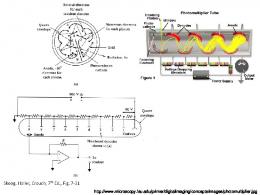
by a membrane containing an aqueous fluid called stroma [17]. ...... To scale up this promising technology, many modules and bag replacements to reduce ...
11 Photobioreactors J.-S. Chang1, *, P.-L. Show2, T.-C. Ling3, C.-Y. Chen1, S.-H. Ho4, C.-H. Tan2, D. Nagarajan1, W.-N. Phong3 1
N A T I O NA L CH E N G K U N G U N I V E RS I T Y , T AI N A N , T AI W A N; 2 UNIVERSITY OF NOTTINGHAM MALAYSIA CAMPUS, SEMENYIH, SELANGOR DARUL EHSAN, MALAYSIA; 3 INSTITUTE OF BIOLOGICAL SCIENCES, UNIVERSITY O F M ALAYA, KUALA LUMPUR, MALAYSIA; 4 H A R BIN INSTITUTE O F T ECHNOLOGY, HARBIN, CHINA
11.1 Introduction to Photobioreactors Phototrophic microorganisms are light-harvesting organisms in which light energy is converted into chemical energy and stored in various forms of carbon biocompounds. Most phototrophic microorganisms are also autotrophic, fixing atmospheric carbon dioxide (CO2) via photosynthesis. The most popular phototrophic microorganisms of commercial interest are cyanobacteria (also known as blue-green algae) and microalgae [1]. Their growth requires light energy and an inorganic carbon source (usually CO2). Therefore, the design of effective PBRs that allow sufficient light penetration and an efficient supply of carbon sources is crucial to the cultivation of microalgae and their further commercial applications. Microalgae are considered third-generation feedstock for producing biofuels and biobased chemicals [2]. Compared with using first-generation (oil palm, soybean, rapeseed, and sugar cane) and second-generation biofuels (lignocellulosic biomass, and agricultural wastes) [3], generating biofuels from microalgae has several benefits: (1) high oil content (ranging from 20% to 70%) of oleaginous algae, (2) higher yields per unit area of land, (3) the ability to use nonarable land for algal cultivation, and (4) cultivation of microalgae in nonpotable water such as wastewater and flue gas [4]. Microalgae also have the ability to produce a wide range of biofuels, including biodiesel [5], bioethanol [6], biohydrogen [7], syngas [8], biobutanol, [9] and bioelectricity [10]. Industrialization of microalgae products requires large-scale culturing, which is commonly done using open systems (open or raceway ponds) or closed systems (PBRs). Although open systems are economic in terms of construction, operation, and maintenance, some major problems include contamination, evaporation, susceptibility to weather conditions, and extensive land requirements (shallow open ponds are generally used to allow good sunlight penetration). In contrast, closed PBR systems can eliminate *
Corresponding Author.
Current Developments in Biotechnology and Bioengineering: Bioprocesses, Bioreactors and Controls http://dx.doi.org/10.1016/B978-0-444-63663-8.00011-2 Copyright © 2017 Elsevier B.V. All rights reserved.
313
314
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
contamination and evaporation issues and achieve higher biomass concentrations. Nevertheless, PBRs have a high capital cost (it is expensive to build PBRs), difficulty in scaling up, and high shear stress (resulting in cell damage) [4,11]. However, because PBRs offer high yields and control of various culture parameters, it is worth investing in developing efficient, low-cost PBR designs and improving PBR operations to reduce the deleterious shear stress in cultures. Despite enormous research, phycologists have yet to come up with a foolproof PBR design that can be used universally for large-scale culturing of microalgae. This is mainly because the choice of PBR depends heavily on the microalga species, desired products, and yield [12]. The most important factor for PBR design is maximizing light penetration into the reactor. This implies a high surface-to-volume ratio and the use of transparent building materials such as acrylic. If a lower surface-to-volume ratio design is preferred, internal LED lighting can improve light penetration and distribution. LEDs are preferred over fluorescent lights because they have lower heat waste loss and are more energy efficient. The second critical factor in PBR design is gaseous mixing within the culture media. Mixing can be achieved either physically (stirrers) or pneumatically (air spargers). The two most common configurations for air spargers are: (1) injecting air at certain points in the system (riser tube) [13] or bubble air from the bottom of the reactor; and (2) passing the culture media through a gas exchanger [14]. Apart from sparging, degassing dissolved oxygen from the culture media is also crucial because it prevents toxic buildup of oxygen in the medium. When the concentration of dissolved oxygen is above air saturation, photosynthesis is inhibited in most microalgae [15]. In this chapter, we discuss major factors affecting the efficiency of PBRs, such as light utilization, mixing, CO2 assimilation, and O2 removal, taking microalgae as target autotrophic organisms for cultivation. The advantages and disadvantages of both open systems and closed PBRs for the cultivation of microalgae are compared and criteria for selecting an appropriate PBR are presented. Finally, current advances in PBR technology in various commercial applications are reviewed.
11.2 Major Factors Affecting Photobioreactor Efficiency Although the growth rate and maximum biomass concentration of phototrophic microorganisms vary with species, most reports agree that microalgae, like many other microorganisms, could grow much faster under favorable environmental conditions. This indicates that optimizing the cultivation condition with a suitable PBR to increase biomass productivity will reduce overall production costs. To enhance the economic feasibility of using phototrophic microorganisms for downstream applications, a better understanding of both the physiological requirements of microalgae and the engineering concepts behind successful PBR design is essential. Several major factors that influence the efficiency of PBR operations are described in detail subsequently. Of these, sunlight (or an artificial light source) and CO2 are direct participants in photosynthesis and hence significantly affect the cell growth and biomass production of microalgae. Any variation
Chapter 11 � Photobioreactors 315
from optimum conditions severely affects photosynthetic efficiency. Other factors such as pH and temperature indirectly affects growth by altering the optimum conditions for cellular functions, which include photosynthesis.
11.2.1
Light Utilization
The most important determinant of photosynthetic efficiency is the availability of light in the required intensity and spectral quality. Through photosynthesis, phototrophic microorganisms can convert light energy from sunlight into chemical energy in the form of carbohydrates [16,17]. Photosynthesis takes place in chloroplasts, which are enclosed by a membrane containing an aqueous fluid called stroma [17]. In the photoactive sites of photosystem I and photosystem II, sunlight can be harvested by the antenna chlorophylls and transferred to the special chlorophyll dimer (P680 and P700) in the reaction center. Excitation of this chlorophyll dimer initiates the electron transfer across membranes and generation of adenosine triphosphate [18]. Cell growth of phototrophic microorganisms is often strongly affected by the level of light intensity, which can fall into one of three categories: light limitation, light saturation, or light inhibition [19]. Below the saturated light intensity, light is one of the key limiting factors for biomass production and photosynthesis efficiency increases with an increase in light intensity. Algae adapt to light intensity variations by a process called photo-adaptation/photoacclimation by changes in cell organization, and increase photosynthetic efficiency. At saturation of light intensity, the rate of photon absorption exceeds the rate of electron turnover and there is no further increase in photosynthesis. With a further increase in intensity, photo-inhibition occurs, causing irreversible damage to the photosynthetic apparatus and inducing oxidative damage to the cells [20]. Because of all these, the influence of light on photosynthetic efficiency also depends on the individual strain and needs to be determined individually. For instance, Ho et al. showed that the biomass productivity of microalgae Scenedesmus obliquus CNW-N increased threefold with an increase in light intensity from 60 to 420 mmol/m2/s [19]. Sun et al. [21] also observed an increase in the biomass concentration of microalgae Neochloris oleoabudans HK-129 from 1.2 to 1.7 g/L when the light intensity was increased from 50 to 200 mmol/m2/s. However, light supply is different from other environmental factors in the sense that its level cannot be uniformly kept inside the PBRs. For example, in the real case of outdoor cultivation, sunlight is often too intense for cells in the surface, resulting in photoinhibition. On the contrary, light intensity at the depth of cell suspension is significantly reduced because most light has been absorbed or scattered by the outermost layer of cells, resulting in light limitation [22,23]. In higher biomass concentrations, light attenuation occurs. The incident light is absorbed by the cells on the surface, creating a shading effect. This also causes two different growth states of algae in the reactor: a photic zone (high light intensity) where photosynthetic growth occurs and a dark zone (low light intensity) where fermentative growth occurs. Efficient mixing ensures that the cells experience alternative lightedark cycles and increase biomass productivity and PBR
316
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
efficiency [24]. In addition, only the spectrum of sunlight from 400 to 700 nm is used by phototrophic microorganisms as an energy source, known as photosynthetically active radiation (PAR) [17]. It was found that growth of many phototrophic microorganisms (e.g., microalgae) under red light (600e700 nm) and blue light (400e500 nm) could induce photosystems, meaning that red and blue lights seems more suitable for cell growth [25]. Thus, increasing the use of PAR is highly correlated with enhancing cell growth of phototrophic microorganisms. This can be done by designing the PBR with a higher surface-to-volume ratio and/or shorter light penetration path to obtain a higher light-receiving area for phototrophic cells [22]. Ho et al. [26] have shown that 1.4 times greater biomass productivity of microalgae Chlamydomonas sp. JSC4 was obtained by a slim-type tubular PBR with a high surface-to-volume ratio of 80.1 m2/m3.
11.2.2
pH
The pH value is an important factor in microalgae cultivation, affecting many biological mechanisms associated with cell growth, metabolism, and uptake of ions [27]. Growth of microalgae can be significantly affected by changes in pH, because pH determines the solubility and availability of CO2 and other essential nutrients, activities of intracellular and cell walleassociated enzymes, and production of various commercially important products. Acidic pH alters nutrient uptake capabilities, interferes with cellular processes, and induces metal toxicity. Alkaline pH lowers the affinity of algae for CO2 and increases the flexibility of mother cells, delaying completion of the cell cycle [28]. The photosynthetic assimilation of carbon has the potential to increase extracellular pH, and release of CO2 by dark respiration can decrease pH. Deviations from optimum pH might inhibit photosynthesis owing to the sensitivity of the photosynthetic apparatus, and the enzymes do not operate well in extreme pH ranges [29]. Previous study demonstrated that the principal carbon fixation enzyme RuBP carboxylase is functional only in the internal pH range of 6.5e8.5. The extreme variation in external pH also influences internal pH and affects photosynthetic efficiency. The external pH also determines the available species of dissolved inorganic carbon (DIC) in the medium. Below pH 7, CO2 is the predominant form of DIC with a minimum amount of HCO3 present. At pH 7e10, HCO3 is the predominant form (about 90%), and above pH 8, CO3 2 is the most available form [30]. The suitable pH for most phototrophic microorganisms varies with species, approximately in the range of 6e10. Optimum pH for any isolated strain is the pH value in which the strain is adapted to grow, and any change may affect growth and biomass concentration [30]. Also, most microalgae (e.g., green algae and diatoms) typically prefer to grow in the pH range from neutral to weak alkalinity [31]. Microalgae growing in neutral to near alkaline pH levels can efficiently accumulate HCO3 into their cells [32]. Freshwater microalgae have been known to tolerate a wide range of pH owing to the fluctuating pH range in the water bodies, whereas marine microalgae have little tolerance to changes in pH, because the pH of seawater is maintained fairly well at around 8.2 [30,32]. For example, Breuer et al. [33] reported that the optimal pH for the
Chapter 11 � Photobioreactors 317
growth of S. obliquus UTEX 393 is in the range of 6e8. Khalil et al. [27] showed that the optimal pH for Chlorella ellipsoidea is about 10. However, in the case of using pilotscale PBRs to decrease the cultivation cost, flue gas is often used as the main carbon source for cell growth and the flue gas may significantly affect the pH balance during the cultivation process [34]. In general, flue gas is mainly composed of CO2, NOx, and SOx [34]. Increasing CO2 concentration would drop the pH to 5, and if the concentration of SOx increased, the pH would decrease to even lower than pH 2e3. This pH change will dramatically damage the cells [35,36]. This indicates that inhibition of cell growth is often caused by pH changes during cultivation, which can be prevented via preparation of buffered medium or installation of a pH-stat system coupled with PBRs.
11.2.3
Temperature
Temperature is one of the most important factors in the cultivation of microorganisms and is highly correlated with the growth rate [37]. The effect of temperature on microalgal growth represents a typical bell-shaped curve in which the growth rates increases with an increase in temperature until the optimum temperature is reached and there is a sharp decline thereafter [6]. Suboptimal temperatures can affect the viscosity of the cytoplasm, leading to inefficient carbon and nitrogen use. Low temperature can also lead to photo-inhibition [28]. Lower temperatures can increase unsaturated fatty acid content to overcome the rigidity of the cell wall. Increased temperatures reduce protein synthesis or denature proteins, inhibiting cellular metabolism. Increased temperatures also cause photo-oxidative damage, and microalgae respond by increasing the pigment concentration as an antioxidant measure. Dunaliella salina produced increased amounts of b-carotene when grown under high irradiance and a high temperature of 35� C [38]. The production of astaxanthin by Haematococcus pluvilais can also be induced by increasing the temperature to 30� C, whereas the optimum temperature for growth is 20� C [39]. Many microalgae can adapt to a temperature drop of up to 15� C below optimum, but an increase of 2e4� C above optimum temperature can adversely affect cell growth and cause cell death. For instance, the specific growth rate of Chlamydomonas reinhardtii BAF-J5 increases with temperature over the range of 17e32� C [40]. Xin et al. [41] also reported that the biomass concentration of Scenedesmus sp. LX1 increases with an increase in temperature from 10� C to 25� C but then decreases with a further increase in temperature to 30� C. It must also be considered that under laboratory conditions, temperature is always measured and maintained, but outdoor culture systems experience seasonal and diurnal variations in temperature [42]. The tolerance of phototrophic microorganisms to ambient temperature needs to be intensively considered with regard to outdoor cultivation, because this may significantly influence cell growth [43]. Ho et al. [6] showed that S. obliquus CNW-N can grow well on most days in subtropical areas, except for a few hot or cold days in a year (i.e., over 33� C or under 18� C). In addition, some studies reported the strong inhibition of growth of phototrophic microorganisms
318
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
by sudden heat stress from flue gas, which is often emitted from power, coal, or steel plants at more than 120e150� C [44]. Selecting a thermophilic strain with a good heatremoval PBR system is important, which can be cost-effective than an expensive heatexchanger system [34]. Few strains have been identified which can tolerate high temperature up to 50� C [45]. When using the same culture conditions outdoors, the temperature of the culture medium would vary with the type of PBR used. It is also shown that overheating can occur on especially hot days, when the internal temperature in the PBR might reach as high as 55� C [11]. For instance, although the closed flat-plate PBR represents a larger light-receiving area with better light distribution, the excess irradiance sometimes may transform into fluorescence and cause heat to be accumulated inside the PBR, resulting in a temperature increase.
11.2.4
Carbon Dioxide Assimilation
Carbon, the major constituent of microalgal cells, which account for more than 50% of the cell mass, is obtained in the form of CO2 for photoautotrophic algae. The atmospheric concentration of CO2 is low (0.03e0.04%) and not optimal for efficient photosynthesis [16]. Also, most phototrophic microorganisms (e.g., microalgae) have low CO2-binding capacity of rubisco (ribulose 1,5-bisphosphate carboxylase/oxygenase), which is the key enzyme of CO2 assimilation through the Calvin cycle [46]. Microalgae have adapted to this problem by developing carbon concentrating mechanisms (CCMs), in which external available carbon is efficiently accumulated inside the cell. This internal carbon pool feeds efficiently into photosynthesis, overcoming the low CO2 concentration available. Wang et al. [47] exploited the CCM mechanism of algae and with fed-batch cultivation C. reinhardtii accumulated 71% of carbohydrates in the cell as reserve. It has been reported that CO2 accumulation by CCM is highly correlated with the CO2 feeding concentration [48]. This suggests that the cell growth of phototrophic microorganisms could be highly influenced by adjusting the feeding CO2 concentration. For large-scale cultivation, capturing the atmospheric CO2 through gaseliquid exchange from air to microorganism is the most commonly used method [49]. Because the atmospheric CO2 concentration is low, capturing CO2 from flue gas containing 6e20% CO2 seems to be a reasonable solution to obtain a better biomass concentration. However, the high CO2 concentration may lead to a drop in pH, which inhibits cell growth. Several phototrophic microorganisms have shown high CO2 tolerance (in the range of 40e100%) [16,34], which is beneficial for the CO2 fixation process of microalgae when using flue gas as a source of CO2. Apart from CO2 concentration, the flow rate of inlet gas also has an important role in determining total inorganic carbon dissolved in the liquid phase [50]. Overall, increasing the CO2 transfer rate from the gas to liquid phase is necessary to enhance the “dissolved CO2” concentration, which highly affects the cell growth of phototrophic microorganisms [50]. Several reports have proposed increasing the CO2 transfer rate by improving mixing, gas injections, or gas recirculation [51]. A well-designed PBR usually should have a larger light-receiving
Chapter 11 � Photobioreactors 319
area and advanced CO2 diffuser allowing a longer CO2 retention time, which can significantly enhance the penetration of light and the dissolution of inorganic carbons [34].
11.2.5
Oxygen Removal
From the mechanisms of photosynthesis, it has been reported that the water-splitting activity of photosystem II is responsible for oxygen generation [46]. Because most phototrophic microorganisms (e.g., microalgae) have lower CO2 binding activity than O2 in rubisco enzymes, the excess O2 apparently competes with CO2 for rubisco binding sites, leading to a switch from photosynthesis to photorespiration, thus significantly reducing photosynthetic efficiency [22]. The presence of excess oxygen, along with excess light, can create reactive oxygen species, causing oxidative damage to the cells. As a rule of thumb, the dissolved O2 concentration in the medium should not exceed 400% of the air saturation level, and algae cannot withstand increased levels of CO2 for more than 2 or 3 h [42]. This kind of oxygen inhibition of photosynthesis is discussed as the Warburg effect or photorespiration. Overall, the excess O2 accumulation is a serious factor that decreases the efficiency of PBRs. In general, O2 removal is one of the most difficult challenges to overcome in designing effective PBRs, especially closed PBRs (e.g., tubular PBRs) [22]. For a closed PBR system, the problem of O2 accumulation increases with an increase in the working volume of PBRs [23]. Hence, it may be necessary to install a separate aeration or degassing system with large-scale closed PBRs to remove excess O2 during cultivation.
11.2.6
Agitation/Mixing
Agitation or mixing is an important feature of any bioreactor. In fact, Suh and Lee [42] stated that when other factors are nonlimiting for algal growth, efficient mixing is the most important factor for high yields of biomass. Mixing ensures the uniform distribution of nutrients to the cells and efficient light utilization, facilitates heat transfer, promotes gas exchange, and maintains the cells in suspension. In PBRs, mixing is also important because it moves cells from the photic to the dark zone, thereby artificially providing lightedark cycles to enhance productivity. In general, the agitation system in PBRs can be divided into the nonmechanical type (e.g., air-lift, bubble column, and tubular PBR) and mechanical type (stirred tank and raceway pond) [23]. However, turbulence created by these methods can elicit hydrodynamic stress, resulting in cell breakage and the inhibition of metabolic activity. The tolerance of microalgae to turbulence is strain dependent and needs to be evaluated individually [11]. Through proper agitation, most phototrophic microorganisms and nutrients could be uniformly distributed in the medium, which is beneficial for cell growth [23]. Appropriate mixing in PBR also helps provide better light distribution over cells [23]. In addition, efficient gas exchange is always associated with proper agitation during cultivation, which can provide better mass transfer of CO2 to cells as well as the effective removal of oxygen gas and
320
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
accumulated heat. All of these clearly show that appropriate agitation is one of the most important factors affecting the efficiency of PBRs.
11.3 Types of Photobioreactors Phototrophic microalgae are used to produce various types of metabolites, antioxidants, polysaccharides, vitamins, bioactive compounds, and polyunsaturated fatty acids(PUFAs), which are important in pharmaceutical, cosmetic, food, and aquaculture industries [52]. The most common phototrophic cultivation condition for microalgae growth involves the use of sunlight as the energy source and carbon dioxide as the carbon source, converting it to biomolecules and concomitantly producing oxygen through photosynthesis [53,54]. An effective PBR is required for the mass production of microalgal biomass with different compositions and functionalities. Typically PBRs consist of four-phase systems: cells as solid phase, growth medium as liquid phase, gaseous phase, and light radiation field [24]. When considering the design of PBRs, issues such as gaseous exchange, mixing pattern, contamination control, light supply efficiency, building material, and geometrical configuration must be considered [12]. Furthermore, the desired end product in the cultivated biomass and the quality of the target products are key factors in determining the design of PBRs [55]. PBRs are classified based on their design [55]; the main PBRs that are developed can be broadly divided into two major types [56]: open systems (such as circular ponds, raceway ponds, and so on) and closed systems (such as tubular PBRs, flat-plate PBRs, internally illuminated PBRs, heterotrophic fermenters, and so on). The types of PBRs are discussed in detailed as follows. The advantages and disadvantages of open and closed PBRs are described in Table 11.1.
11.3.1
Open-Type Photobioreactors
In open-type PBRs, the culture is directly exposed to the environment [57]. They have traditionally been used to cultivate microalgae because to their low cost, simplicity [52,55], and low energy consumption, and the high availability of solar radiation [55]. The most commonly used open systems are circular and raceway ponds. Open systems are still widely employed in industrial processing, but they have little room for further technological improvement. This is because of various constraints related to their operation for large-scale applications [12]. Drawbacks of open systems include easy contamination, limited control of culture conditions [55], evaporative water losses, temperature fluctuations, inefficient homogenization resulting in poor mass transfer rates, nutrient limitation, and uneven light intensity [4]. There are also other major concerns associated with open cultivation systems, such as poor control over environment parameters caused by the direct influence of weather conditions and geographical locations; the requirement of large areas of land to construct shallow culture ponds; and high harvesting costs owing to low biomass yield [4,12]. Because
Chapter 11 � Photobioreactors 321
Table 11.1 Comparison of Open and Closed Systems for the Cultivation of Microalgae [11,12,64,134] Feature/Parameter
Open Systems
Closed Systems
Space occupied Surface area to volume ratio Variability of cultivable microalgal strain Main criteria for strain selection Products range Cultivation period and growing season
Large Low Restricted Growth competition Low-value products Limited, highly dependent on location and weather Low Low High Difficult None High Very low Mixing Poor Low High Poor Low Poor Absolute Difficult Simple Almost impossible Impossible Low Low Low Low Not susceptible Less possible Low Easy Low Low Small
Small High Extensive and flexible Shear resistance High-value products Extendable
Microalgal cell density Harvesting efficiency Risk of contamination Contamination control Sterility Water loss Hydrodynamic stress on microalgae Most expensive parameters Mixing Gas transfer control Carbon dioxide losses Removal of oxygen Super dissolved oxygen concentrations Efficient use of light Weather dependence Temperature control Process control Flexibility of production Standardization Overheating problems Biomass concentrations during production Biomass concentrations at harvesting Productivity Quality of biomass Reproducibility of production Treatment process efficiency Maintenance Construction costs Operation costs Capital investment
High High Low Easy Achievable Preventable Low to high Temperature and oxygen control Uniform High Preventable Efficient High High Insignificant More uniform Complicated Possible Possible High High High High Susceptible Possible High Difficult High High Big
open culture systems are still widely used in the commercial production of microalgae because of economic concerns, it is still necessary to choose an appropriate open-type culture system with suitable operating strategies based on the characteristics of the cultivated microalgal species.
322
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
11.3.1.1 Circular Ponds With the principal advantage of using abundant sunlight as the energy source for microalgal growth, circular ponds are one of the oldest pond types used for commercial cultivation of microalgae such as Chlorella sp. [58]. These ponds are constructed of concrete and are equipped with a rotating arm mounted at the center to mix the culture (Fig. 11.1). They are also lined with materials such as plastic sheets or inert membranes as a secondary containment measure. As the rotating arm travels over a greater distance, greater mixing of the culture can be attained at the periphery of the ponds [57]. Thus, circular ponds are most suitable for growing microalgae that easily precipitate, such as Chlorella vulgaris and Chlamydomonas sp.
11.3.1.2 Raceway Ponds Raceway ponds are the most widespread reactors used to culture microalgae commercially owing to their flexibility and ease of scaling up [55]. In raceway ponds, microalgal cells, nutrients, and water are continuously circulated around a racetrack (Fig. 11.2). The depth and flow rates of the raceway ponds affect the distribution of nutrients. The paddlewheel provides circulation and mixes the nutrients and cells, and also prevents sedimentation. Microalgae are kept suspended in the water and are recirculated to the surface on a regular frequency, following the driving force of liquid flow generated from the paddlewheel [59]. Usually, raceway ponds are operated in a continuous mode by adding fresh culture medium at a position before the paddlewheel. After completion of the circulation loop, microalgal culture is then harvested behind the paddlewheel [59,60]. Bubbling carbon dioxide can enhance aeration, carbon dioxide consumption, and mixing, and thereby increase the growth of microalgal cells [60].
FIGURE 11.1 Circular open pond (300-ton capacity) located at the An-Nan campus, National Cheng Kung University, Tainan, Taiwan.
Chapter 11 � Photobioreactors 323
FIGURE 11.2 Four-channel raceway pond (20-ton capacity) located at the An-Nan campus, National Cheng Kung University, Tainan, Taiwan.
Because sunlight can penetrate pond water only to a limited depth, raceway ponds are usually shallow (30e50 cm) to ensure sufficient light supply [59]. Raceway ponds are cost efficient and energy efficient because of the low construction cost, low energy requirement for mixing, and easy control of culture depth and liquid movement [55,61]. However, their performance is constrained by the adaptability of microalgae strains, land area requirement, low efficiency for light use, poor gas diffusivity owing to the limited contact time between gas and culture, high risk of contamination, poor temperature control, and low biomass density [12,13,24,59]. Not many contaminants can survive under extreme conditions; thus, contamination can be avoided by cultivating some highly resistant microalgal strains at high pH or high salinity in raceway ponds. The most successful commercial cultivation of algae in raceway ponds includes Chlorella sp., Spirulina sp., and Dunaliella sp., which are cultivated under stringent conditions that inhibit the growth of other weed algae or pathogenic bacteria. However, pH gradients resulting from inefficient mixing system can cause contamination in those cultures [62]. To overcome the problem of poor mass transfer efficiency, raceway ponds are frequently equipped with sumps or mixing columns to increase gas and liquid contact time [63,64].
11.3.1.3 Multilayered Photobioreactors The spatial distribution of light in a PBR is crucial and the light-shading effect could be a factor limiting the microalgal production rate, especially at higher cell densities [65]. A light-shading effect can be defined as a phenomenon in which the high density of microalgal cells in a PBR reduces the penetration of light into the culture broth, limiting
324
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
Medium Layer Algae Layer
c Partition Layer
Light
FIGURE 11.3 Basic structure of a multilayer PBR.
the photosynthetic ability of cells near the center or bottom of the PBR [65e67]. However, light distribution can be improved by using a multilayered PBR. The structure of the interior layered compartments enables the dilution and uniform diffusion of a strong incident light in a reactor [65]. In multilayered PBR (Fig. 11.3), high-density microalgal cells are immobilized and fixed onto artificial support materials such as filter paper (pore 0.45 mm), nylon mesh, cotton rope, a polyethylene screen, polystyrene foam, and a concrete layer to form a thin layer of algal biofilm [68]. A clear partition is furnished with a semipermeable membrane layer to hold the culture medium for microalgae growth. Multiple layers of algal biofilms and clear medium layers are arranged alternately in an array style [65,69]. Gaskets are used to prevent the leakage of cell suspension, medium, or produced gas. Substrates and nutrients transfer between the algae biofilm and medium layer, but bacterial cells are restrained from migrating between them owing to the small pore size of the permeable membranes. The clear medium layers enable high light transmittance to be maintained on both sides of the algae biofilms. Along the clear medium layers, incident light is induced and diffused evenly into the PBR [65]. With the objective of suppressing the light-shading effect, Kondo et al. [65] constructed a multilayered PBR to investigate hydrogen production efficiency by photosynthetic bacteria. Compared with a conventional plate-type reactor, more light energy is delivered to the interior of the multilayered PBR, which may contribute to an increase in photosynthetic activity. The study successfully showed that hydrogen productivity in the multilayered PBR increased twofold compared with a plate-type reactor. Furthermore, an experiment carried out by Cheng et al. [70] showed that the high biomass productivity of green alga Botryococcus braunii could be attributable to improved light availability in the multilayer arrayed PBR. This finding was also in agreement with the work of Liu et al. [69], in which proper diffusion of strong incident light in PBRs boosted microalgal biomass production efficiently. Outdoor cultivation of the oleaginous microalga S. obliquus using this cultivation method showed 400e700% higher biomass productivity over a conventional open pond. In addition to providing
Chapter 11 � Photobioreactors 325
excellent light penetration efficiency, this attached method also offers a few other possible advantages over conventional open ponds, such as contamination control, water saving, and easy harvesting and scale-up [69].
11.3.2
Closed-Type Photobioreactors
Current research on PBRs has focused mostly on closed systems. The culture is entirely enclosed within the PBR [57], with no direct gaseous exchange with the atmosphere [55]; it allows sunlight or artificial illumination where phototrophic microalgal growth can proceed without light limitations [53]. Closed systems are designed to overcome the constraints of open systems. Unlike open raceway ponds, closed PBRs have two different areas that involve in light utilization efficiency. The first area receives direct light and the second area collects diffuse light [55]. Carvalho et al. [12] suggested that various configurations of closed systems are appropriate for higheshear sensitive microalgal strains or final products that are highly susceptible to microbial degradation. In general, close systems are advantageous over open systems because they: 1. are flexible in controlling and maintaining optimized reproducible cultivation conditions such as temperature, light, pH, and carbon dioxide concentration [59] 2. allow better control of contaminants [52] 3. prevent water evaporation [59] 4. minimize carbon dioxide loss [59] 5. allow good heat transfer [71] 6. achieve higher biomass yield [59] 7. are able to produce high-quality complex bioproducts by allowing single-strain cultivation of targeted microalgae with lower risk of contamination [59] Despite the ability to produce a wide variety of microalgal strains [55] at higher cell mass productivities [12], the functions of closed systems are limited to the production of high-value microalgal biomass because of higher operation and construction costs compared with open systems for commercial application [55]. Light and carbon dioxide supply are key processing parameters to be considered for improving the efficiency of a closed PBR. The control of nutrient supply, pH, and temperature control are simple and no major improvements are expected [12]. Other factors need to be considered in designing close systems [59]: 1. The closed PBR should allow the cultivation of various microalgal strains universally. 2. The closed PBR should provide uniform illumination of the culture surface and rapid mass transfer of carbon dioxide and oxygen. 3. The closed PBR should minimize fouling of the light-transmitting surfaces of PBRs. 4. The closed PBR should allow mass transfer at high rates to avoid suppression of cells.
326
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
5. The closed PBR should be able to operate under intense foaming conditions caused by high rates of mass transfer. 6. The closed PBR should display a high illuminated ratio of surface area to volume (A/V) and ensure maximum light illumination to prevent the light-shading effect. Among all of these factors, the provision for light penetration influenced by the surface-to-volume ratio is the main parameter that affects reactor design. To achieve a high ratio of surface to volume, several types of PBR have been developed [12]. For example, tubular and flat-plate reactors are the most popular choice [72] and are specifically designed for efficient harvesting of sunlight. On the other hand, the fermentertype PBR is designed for artificial illumination [12].
11.3.2.1 Tubular Photobioreactors Tubular PBRs such as stirred tank, bubble column, and air-lift PBRs are currently the most widely used closed systems on an industrial scale [52,55]. They allow better control of culture conditions and reduce the risk of contamination [55]. In tubular PBRs, microalgal culture flows through solar collector tubing and is recirculated by mechanical pumps or aeration [55]. For example, the bubble column and air-lift are usually composed of vertical tubing with high transparency to allow good light penetration and enable a sufficient supply of carbon dioxide via bubbling [12]. Furthermore, good overall mixing and efficient removal of excess dissolved oxygen can also be achieved by bubbling air at the bottom [12]. In most cases, bubbling can be done via air pumps and usually in the plug flow direction [62]. To prevent the cells from stagnating in the dark interior of the reactor tubes or at the bottom of the reactor (known as dead space), it is important to ensure that the flow in the tubes is turbulent when designing a tubular PBR [55]. The dimensions of fluid microeddies should exceed those of the microalgal cells to prevent turbulenceassociated damage [55]. Despite potential advantages, the attempt to scale up tubular PBRs for commercial production is challenging and further studies are needed to overcome all of the setbacks. 11.3.2.1.1 STIRRED TANK PHOTOBIOREACTORS Stirred tanks are the most conventional type of PBRs, particularly for the production of high-value products such as fine chemicals or pharmaceuticals [11]. Stirred tank PBRs (Fig. 11.4) ensure precise control over processing parameters and maintain the sterility of microalgal cultures. These advantages are crucial for the production of high-value products [58]. Agitation is provided mechanically using impellers of different shapes and sizes. Vortex can be reduced by using baffles. The carbon source for microalgal growth is supplied by bubbling carbon dioxideeenriched air from the bottom of the reactor [59]. However, stirred tank PBRs have a few limitations. Illumination provided externally using LEDs, optical fibers, or fluorescent lamps cannot reach maximum microalgal photosynthetic efficiency. This is because of the low ratio of surface A/V in the reactor
Chapter 11 � Photobioreactors 327
Impeller
Sparger
Air + CO2
FIGURE 11.4 Basic structure of a stirred tank PBR.
system and an insufficient provision of light in terms of cost and energy. This can be overcome to some extent by efficient stirring mechanisms. The unused portion of sparged gas and oxygen produced during photosynthesis diffuses out of the culture and collects in the large disengagement zone in the reactor [59]. 11.3.2.1.2 VERTICAL TUBULAR PHOTOBIOREACTORS Vertical tubular PBRs are compact, user-friendly bioreactors with a high ratio of surface A/V [73], low contamination risk, and high biomass productivity [56]; therefore, they are suitable for large-scale cultivation of microalgae. Vertical tubular PBRs consist of transparent vertical tubes to allow maximum light penetration [23]. They offer efficient mixing with high gas transfer rates and have excellent control over growth conditions. Bubbling of gas from the bottom of the column enables an efficient supply of carbon
328
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
dioxide and optimal removal of excess dissolved oxygen [74]. In this type of reactor, constant agitation of the medium is generated by bubbling carbon dioxide via sparging [56], causing smaller shear stress than with pumps or impellers [73]. The sparger is located at the bottom of the reactor, where the sparged gas is converted into tiny bubbles [59]. In tubular PBR, light availability is affected by the aeration rate, gas holdup, and liquid velocity [75]. Both the height and diameter of a single vertical tubular PBR should not exceed a certain limit, to reduce the problems of mutual shading and light limitations, respectively. For example, an experiment carried out by Sa´nchez Miro´n et al. [15] using a 0.2-m-diameter bubble column PBR with variable height demonstrated that volumetric biomass productivity and eicosapentaenoic acid (EPA) production declined when the column height was more than 5 m. Hence, it was suggested that the optimal dimensions of a bubble column PBR should be approximately 0.2 m in diameter and 4 m high. The aeration rate in tubular PBRs should be adjusted within acceptable range by considering factors such as sensitivity to shear stress [76] and light penetration [15]. The aeration rate should be kept to a certain minimal level to prevent cell stagnation for long periods in the dimly lit interior of the PBRs. In contrast, high aeration rates could reduce light penetration over time by generating an accumulation of persistent microbubbles [15]. Bubble column and air-lift PBRs fall under this category and can be differentiated based on the mode of liquid flow [23]. Air-lift PBRs possess good mixing properties whereas the configuration of bubble columns allows efficient aeration [56]. 11.3.2.1.2.1 BUBBLE COLUMN PHOTOBIOREACTORS Bubble column PBRs consist of vertical cylindrical columns and the light source is usually supplied externally [59] (Fig. 11.5). It is advantageous in terms of low cost, a large ratio of surface A/V, good mass and heat transfer, homogeneous culture condition, and efficient removal of oxygen [59]. Efficiency of photosynthesis depends greatly on the gas flow rate. The increase in gas flow rate that leads to shorter lightedark cycles could significantly increase the photosynthetic efficiency of microalgae [59]. Separation of gas and liquid takes place at the freeboard regime that is located at the top of the reactor [71]. Carbon dioxide mass transfer and mixing is done through gas bubbling from a sparger [23]. The mixing pattern in bubble column PBRs is different from that of tubular PBRs. In bubble columns, mixing can be random and erratic via turbulence created by the gas mixture sparging from the bottom of the PBRs [15,71,77]. As a result, microalgal cells may be exposed to uneven light intensity for a long time and cell sedimentation is more likely to occur [78]. For the purpose of scaling up, turbulence can be increased by installing perforated plates inside the column to break up and redistribute coalesced bubbles [71]. 11.3.2.1.2.2 AIR-LIFT PHOTOBIOREACTORS Air-lift PBRs have two interconnecting zones: the riser and the down-comer (Fig. 11.6). The riser is the region where gas mixture is sparged, whereas the down-comer is the region that does not receive gas [23]. In the
Chapter 11 � Photobioreactors 329
FIGURE 11.5 Bubble column PBRs (50-L capacity).
riser, mixing is done by bubbling gas via the sparger and no physical agitation is involved. Similar to bubble columns, sparged gas moves upward randomly in the riser with the aid of gas holdup by the riser. Fluid dynamics is significantly influenced by gas holdup by the down-comer. The degassed liquid moves downward with defined and oriented motion. The difference in gas holdup between the riser and the down-comer is a crucial parameter that should not be overlooked when designing air-lift PBRs [23,59,79]. Common configurations used for air-lift are internal loop, internal loop concentric, and external loop vessels. Of these, the external loop configuration ensures better mixing because of the distance between the riser and the down-comer, which enables efficient gas disengagement. Air-lift PBRs enable a higher growth rate of microalgae compared with bubble column PBRs because of the better mixing pattern [76]. Air-lift PBRs are characterized by their advantage of generating a circular and homogeneous mixing pattern, which can provide flashing light effects to microalgal cells. This happens when the liquid culture moves continuously through dark (riser) and light (down-comer) zones [76,80,81]. Furthermore, the residence time of gas in various zones has a significant influence on a few parameters such as heat transfer, mass transfer, mixing, and turbulence [82].
330
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
Draft Tube
Sparger
Air + CO2
FIGURE 11.6 Basic structure of air-lift PBR.
Air-lift PBRs are generally recommended for microalgal cultures that are fragile and sensitive to shear stress. Efficient mixing can be achieved by increasing the aeration rates, but should be done moderately to prevent cell damage [62]. Oncel and Sukan [83] observed that air-lift PBRs can sustain better biomass yields of different microalgae than bubble column PBRs. This was probably because of better mixing, which can prevent cell sedimentation and improve the efficiency of light utilization in air-lift PBRs [71,83]. Monkonsit et al. [81] also proved that under the same operation conditions for both reactors of the same size, the cultivation of diatom Skeletonema
Chapter 11 � Photobioreactors 331
costatum achieved better biomass productivity (6.4 � 104 cells/s) in the air-lift than the bubble column PBR (2.2 � 104 cells/s). However, in large-scale production, both air-lift and bubble column PBRs have the problem of light limitation. It was suggested that installing immersed optical fibers as an internal light source could increase light penetration efficiency in high-density cultures [84].
11.3.2.2 Flat-Plate Photobioreactors Flat-plate PBRs have a cuboidal shape with a narrow light path [59] (Fig. 11.7) and are the most popular choice among closed PBRs [72]. Flat-plate PBRs are characterized by a high illuminated surface A/V ratio and open gas transfer area [59,71]. These two characteristics make them suitable for the mass production of microalgae in both indoor and outdoor culture systems [76]. Flat-plates are made of transparent materials for maximum use of light energy [62]. The thickness of the plate is the determining factor for the illuminated surface A/V ratio and the length of the light path. The A/V ratio is high when the thickness is minimal [12]. The light source can be diffused and distributed effectively with thin plate and concomitantly lead to higher optimal cell density and biomass yield through higher photosynthetic efficiency [85]. However, the thin plate is usually expensive and difficult to clean, and is easily subjected to temperature fluctuation and light inhibition [71]. Agitation can be provided in two ways. Air is either bubbled through a perforated tube or rotated mechanically using a motor [59]. Baffles can be included to improve mixing efficiency [86]. The open gas disengagement system reduces the need for a dedicated degassing unit [56]. The accumulation of dissolved oxygen concentrations is low in this type of PBR [76]. As for temperature control, water spray or internal heat exchangers are used to cool flat panels [55].
Medium Inlet
Medium Outlet Air + CO2 FIGURE 11.7 Basic structure of a flat-plate PBR.
332
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
However, one major shortcoming of flat-plate PBRs is fouling. This occurs when cells attach to the plastic walls, causing a reduction in light availability and an increased risk of contamination [55]. To scale up this promising technology, many modules and bag replacements to reduce excessive fouling or contamination are required. Although flatplate bioreactors work well for laboratory tests and small-scale purposes, the scaling up for commercial production of microalgae requires more research and effort [55].
11.3.2.3 Internally Illuminated Photobioreactors The use of sunlight as the sole light source for the cultivation of photosynthetic microalgae has its own limitations, such as seasonal and diurnal variations in light intensity owing to weather, seasons, and geographical location. Typically, such cultivation is possible only in a tropical climate and is limited to warmer months in other areas. Ogbonna et al. [87] showed that the growth of microalgae was stunted during prolonged periods of bad weather such as cloudy or rainy days. The light source is insufficient to support microalgal growth and eventually causes a decrease in biomass productivity and biochemical composition of microalgal cells [87]. Apart from that, a reduction in biomass productivity can occur when the outermost layer of cells experiences photo-inhibition owing to excessive exposure to light, or the innermost layer of cells experiences self-shading as the result of insufficient exposure to light and reduced light penetration. Therefore, efficient use of light is the main factor in designing PBRs [76]. This impediment can be addressed by installing an internal illumination device (such as optical fibers or fluorescent lamps) inside the cultivation vessel [62,76]. Carvalho et al. [12] agreed that the installment of an internal illumination device such as optical fiber was able to provide a more homogeneous distribution of light for better photosynthetic efficiency. Optical fibers are installed for solar light collection, transmission, and distribution of light energy more efficiently and evenly inside PBRs [87], increasing average irradiance and the depth to which light reaches inside the PBRs [88]. This type of PBR can also be modified so that the light intensity of solar and artificial light systems can be controlled [87]. The integration of solar and artificial light devices enables a continuous supply of light to PBRs during cloudy weather or at night [62]. Also, internally illuminated PBRs can be heat-sterilized under pressure and therefore reduce the risk of contamination [87]. However, the use of optical fibers in PBR has a few limitations such as capital costs, difficulty in cleaning, and the loss of light energy in transmission in optical fibers. Hence, establishing its practical feasibility for large-scale application using PBRs equipped with optical fiber for internal illumination remains a challenge [71]. LED was also used as an internal light source at a much lower construction cost [89]. An LED-based internally illuminated PBR has the advantage of using light sources at specific wavelengths. This might be beneficial to the production of specific products from microalgae in case target products can be induced by light sources at a specific wavelength [37,90].
Chapter 11 � Photobioreactors 333
11.3.3
Fermenter-Type Photobioreactors
Fermenter-type PBRs (Fig. 11.8) are the least expanded systems [12] which offer open gas exchange and are mainly used for optimization studies [56]. The main advantage of the fermenter-type of PBRs is the ability to monitor and control every operating parameter precisely and accurately [12,56]. However, scaling up is restricted because of the low surface A/V ratio and high capital costs [56]. For large-volume cultivation, illumination can be supplied internally [91] and agitation can be provided using an impeller or magnetic stirrer [56]. Hence, if the productivity can be enhanced, fermenter-type PBRs would probably become a competitive alternative for the production of biochemical products derived from microalgae [12]. The fermenter-type PBR is also well-suited to heterotrophic growth of microalgae, in which the light source is no longer a limiting factor.
11.3.4
Comparison of the Performance of Different Photobioreactors
The pros and cons of these PBRs are listed in Table 11.2. Apparently, research is required to understand the criteria needed to achieve the best performance in PBRs and overcome bottlenecks affecting existing PBRs [52]. Light penetration and distribution, shear stress, mass transfer, scalability, and the physiology of microalgal cells are crucial parameters that need to be considered in the development of PBRs [59]. The design and performance of PBRs need to be further improved to become more competitive and economically feasible [52]. Each specific microalgal strain and the target product require different cultivation conditions that may not be supplied by a single PBR design. Open Acid/Base Antifoam Medium Inlet
Air + CO2
Vent
Level Sensor
pH Dissolved O2
Medium Impeller
Thermocuople
Harvest Outlet Sparger
FIGURE 11.8 Basic structure of a fermenter-type PBR.
334
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
Table 11.2 Comparison of the Performance of Different Types of Photobioreactors in Microalgal Cultivation [4,74,76,89,96] Photobioreactor Type Examples Open systems
Circular ponds Raceway ponds
Closed systems Tubular
Stirred tank
Advantages
Limitations
� � � � �
Economical Easy to clean Easy maintenance Good for mass cultivation Low power consumption
� Limited control of culture conditions � Poor mixing � Poor light and carbon dioxide use � Difficult to grow microalgae for long periods � Low productivity � Limited to a few microalgal species � Easily contaminated � Small illumination surface area � Constant loss of water � Require large area of land � Decrease in illumination surface area upon scale-up
� � � � � � � �
Less costly Suitable for outdoor culture systems High biomass yields High-value products Large illumination surface area High-value products Small illumination surface area Precise control over culture conditions High mass transfer Good mixing Homogeneous shear Low energy consumption Easy to sterilize, low fouling High potential for scalability Readily tempered Reduces photo-inhibition and photo-oxidation Good for immobilization of microalgae High photosynthetic efficiency Best exposure to lightedark cycles Requires least land area disengagement zone separate for gassed liquid and gas phase Relatively cheap Easy to clean Readily tempered Large illumination surface area Suitable for outdoor culture systems Good light path High biomass yields
� � � � � �
� Vertical tubular 1. Bubble column � � 2. Air-lift � � � � � � � � �
Flat-plate
� � � � � � �
pH gradients Oxygen accumulation Fouling Require large land area Photo-inhibition Limited to heterotrophic microalgae
� � � �
Small illumination surface area Sophisticated construction materials Shear stress to cultures Decrease in illumination surface area upon scale-up � Support costs � Modest scalability
� � � � � � �
Difficult to scale up Difficult temperature control Expensive construction materials Some degree of wall growth Shear damage from aeration Low photosynthetic efficiency Subjected to light inhibition easily
Chapter 11 � Photobioreactors 335
Table 11.2 Comparison of the Performance of Different Types of Photobioreactors in Microalgal Cultivation [4,74,76,89,96]dcont’d Photobioreactor Type Examples
Internallyilluminated
Fermenter-type
Advantages
Limitations
� Good for immobilization of microalgae � Low oxygen buildup � Shortest oxygen path � Low power consumption � Allows illumination from inside � Increases average irradiance � Good light penetration � Can be heat-sterilized under pressure � More homogeneous distribution of light � Reduces dayenight cycles and diurnal variation in light intensity � Offers open gas exchange � Mainly used for optimization studies � Monitors and controls every operating parameter precisely
� Subjected to fouling easily
� High capital costs � Difficult to clean � Loss of light energy transmission in optical fiber
� Restrict scale-up owing to low area to volume ratio
pond systems could provide this kind of versatility in the case of the strains to be grown, but they are limited to microalgal strains that are superior to others when cultivated in an open environment by means of their growth conditions. Nevertheless, hybrid PBRs have proved to be a promising technology for the mass production of microalgae compared with single PBRs. Hybrid PBRs are developed by combining two different types of PBRs, in which the disadvantage of one PBR is complemented by the other [59]. Hybrids can be devised by combining different types of PBRs to develop more suitable ones for the mass cultivation of microalgae. The selection of a suitable PBR strongly depends on the purpose of the production facility, the microalgae species, and the desired final products. All these PBRs should not be deemed competing technologies; instead, they complement each other in their pitfalls and can help build a multipurpose PBR that would be foolproof and more flexible [74].
11.4 Criteria for the Selection of Photobioreactors 11.4.1
Type and Quality of the Target Product
What is the target product that you want to produce from microalgae? And what is the desired quality of that bioproduct? Knowing the answers to both questions is the first criterion to selecting a PBR for microalgal cultivation. Different types of PBRs have their
336
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
own characteristics, strengths, and weaknesses. If the targeted product is biofuel, for instance, it is essential to produce lipid/carbohydrate-rich biomass using the least amount of resources and the cheapest processes. Only then can the price of the biofuel be competitive with fossil fuels. In this case, an open system (open pond or raceway) is a good choice because of its simplicity and inexpensive installation [92]. However, if highvalue products are desired, such as the health supplements docosahexaenoic acid (DHA) [93], EPA [94], and phycocyanin [95], pharmaceuticals (hormones), and cosmetics [96], the more expensive closed systems become justified and more preferable [97]. This is because closed systems enable manipulation of culture conditions for the accumulation of specific biomolecules in microalgae. Such high-quality products are expected when it comes to pharmaceuticals, because they have to be approved for human consumption. Closed systems also minimize the risk of contamination and are capable of producing biomass of higher quality compared with open systems [11].
11.4.2
Tolerance of Microalgal Strains
The second most important criterion is determining the adaptability and tolerance of microalgae strains under outdoor conditions and shear forces and oxygen buildup generated by PBRs. In open systems, strains must be able to compete with other bacteria and fungi for nutrients. In the case of cross-contamination, primary microalgal strains must be able to grow rapidly despite the presence of other algal species. In closed systems, microalgal strains have to withstand strong shear forces generated by pumping or sparging. In addition, the strains will face possible oxygen toxicity resulting from excessive buildup of molecular oxygen in the closed PBR during photosynthesis [4]. Therefore, a suitable PBR system should be determined based on the ability and tolerance of microalgal strains used to the environmental factors involved in different types of PBRs.
11.4.3
Scale and Performance Versus Cost
The type of PBR dictates the maximum biomass yield obtained; closed systems produce significantly more biomass at a higher quality compared with open systems. The desired production rate of the microalgae biomass will determine the size of the chosen PBR. Open systems can be scaled up easily and inexpensively to accommodate larger production rates. However, open systems also have a low CO2 transfer efficiency, low biomass productivity, risk of contamination, and evaporation of culture medium [64]. Closed systems, on the other hand, can produce biomass containing a higher content of targeted biomolecules owing to thorough mixing and better CO2 mass transfer. Nevertheless, it is difficult and expensive for the scale-up of closed PBRs [12]. Major issues with large PBRs are that they generate high shear forces during mixing and accumulate excess oxygen more rapidly, causing damage to microalgal cells [4]. As such, a balance between biomass yield and the cost of PBR needs to be properly established.
Chapter 11 � Photobioreactors 337
11.4.4
Availability of Resources
The availability of resources such as land, water, CO2, and light must be taken into consideration when choosing a PBR and determining its final size. For open systems, a large flat land that receives ample sunlight is needed to maintain a shallow culture medium for maximum photosynthetic and mixing efficiency. Proper geological positioning is more important for open systems than closed ones. Closed PBRs require less space because they can be designed to expand vertically [96]. CO2 and water resources accounts for most of the cost in the large-scale cultivation of microalgae [98]. The cultivation of algae using flue gas, seawater, or wastewater can dramatically reduce production costs and is desirable from an economic perspective. Weather has a huge role in the growth rate of microalgae. The warm temperatures and ample sunshine in tropical areas are ideal for culturing microalgae. In countries with changing weather conditions, outdoor cultivation cannot proceed during cold winters or the typhoon season [20]. However, because even tropical areas receive an average of only 6e8 h of sunshine daily, supplementary lighting (fluorescent or LED light) can be provided to allow continued growth of microalgae [99]. The availability of professional labor is vital to monitor and maintain PBRs properly. Required skills for labor to maintain the successful monoculture of algae include knowledge about the growth patterns of microalgae and the ability to operate PBRs. Further training needs to be provided if sophisticated PBRs and equipment are used.
11.5 Current Advances in Photobioreactors in Commercial Applications Market trends and consumers’ growing interest in natural and healthy products have pushed researchers and industries to develop novel products with functional ingredients [100e104]. Microalgae have been recognized as a viable source of functional ingredients with positive health effects because these microorganisms produce bioactive components such as PUFAs, polysaccharides, natural pigments, essential minerals, vitamins, enzymes, and peptides. The extraction and applications of these high-value compounds have been reported and studied [95,100,105e109]. The global microalgal biomass market is estimated at US $5e7 billion, the health sector accounts for US $2 billion, and the aquaculture market is about US $0.7 billion (www.oilgae.com, Report on non-fuel algal products, February 2015). Algal pigments and other high-value products are currently on the market and the launch of new products needs considerable research and scale-up of the specified product. Some algal products (fuels and pigments) have to compete on the market with available synthetic derivatives, whereas some products (fish hatchery feed and hydrocolloids) are derived solely from algae. The conventional open pond cultivation system is the most simple and commonly used one for the cultivation of microalgae for commercial purposes. Based on
338
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
techno-economic analyses, it was predicted that raceway ponds are the most economical way to cultivate microalgae commercially [98]. Costs can be reduced significantly if the culture is coupled with bioremediation (e.g., CO2 capture, wastewater treatment). In an open pond system, key factors in microalgae productivity are the circulation and mixing of microalgae biomass, nutrients supplied, and CO2 gas. The commercial products generated from microalgae grown with varieties of PBR systems are discussed subsequently.
11.5.1
Biofuels
Taking into consideration global energy security and the environment, there is an urgent need to develop a clean and renewable energy source. Among potential energy alternatives, biomass energy, which has the feature of mitigating CO2 emissions, is of most interest worldwide. Microalgae have emerged as a new feedstock for producing biofuels because many microalgal species have the ability to synthesize and accumulate large amounts of neutral oils and lipids (the precursor of biodiesel) or carbohydrates (in the form of starch and cellulose) in cells [19,101,110,111]. Hence, oil- and carbohydrate-rich microalgae can be an ideal source for producing biodiesel or bioethanol via transesterification or microbial fermentation. Large-scale, lipid-rich microalgae cultivation not only reduces carbon dioxide emission but also applies the wastewater resources to develop green energy [112,113]. Microalgae technology has the potential to reduce global warming and provide sustainable resources. The conversion of algal lipids into biodiesel is a primary development, whereas algal residue after lipid extraction can be converted into gaseous biofuels through anaerobic fermentation. Industrializing microalgae culture requires biomass productivity evaluation to be conducted per unit land area. Chisti [20] evaluated that it requires approximately 170 hectares (ha) for a microalgae PBR system to produce 10,000 m3 biodiesel (based on 30% by weight algal lipid without specifying a specific PBR). A PBR system by the Dutch firm Algaelink requires roughly 125 ha. The growth of microalgae in plastic bags conducted by GreenFuel in Arizona requires approximately 217 ha. Consequently, the key to evaluating the amount of land required to produce a certain amount of biofuel relies on the design of the PBR system. Currently, 2 ha of available space can produce a maximum of 200 m3/year biodiesel, which is not enough to meet the requirement for a 5-MW diesel power generator factory. Under the condition of limited space, recycling energy from algal residue and adjusting the ratio of power generation with existing green energy sources for a demonstration plant will become important in the future. With limited life cycle analysis data available for microalgae-based biofuel production, the impact on land use seems to be the main advantage, second only to water use and CO2 capture if industrial wastewater and flue gases are used [114]. To compete with the existing fuel structure and demand, microalgae-based biofuels must be produced cost-effectively. Taking other production costs into consideration, selecting a suitable
Chapter 11 � Photobioreactors 339
strain that has high target product content (lipids for biodiesel and carbohydrates for biohydrogen, bioethanol, and biobutanol) and high biomass productivity is of profound importance. Higher product yield and biomass concentration reduce harvesting and biomass concentration costs. In this direction, hundreds of microalgal species are studied by academic researchers and industrialists to come up with a strain that suits production needs. Microalgal species rich in lipid content and mostly exploited for biodiesel production include B. braunii (high hydrocarbon content and oil is secreted in the cell wall), Chlorella sp., Nannochloropsis sp., Tetraselmis sp., Scenedesmus sp., and Dunaliella sp. [97]. Among these, various strains have been used industrially to produce microalgal feedstock. Several companies use genetically modified algae to fulfill the needs of the production process: high lipid yield, salt tolerance for improved outdoor cultivation, and so on. Strains used for microalgae-based fuel production and cultivation methods used are listed in Table 11.3. Of microalgal cultivation methods used for biofuel production, closed systems account for about 52%; the rest is shared by open ponds and natural settings [114]. Mostly PBRs are custom designed for specific purposes, which gives high yields necessary for further treatment processes (Table 11.3). Such custom-made PBRs provide the advantages of low production costs, reduced water use, and capture of waste gases from industries, and they are user friendly. Microalgal biomass residues can be used after oil extraction as a base for anaerobic fermentation of further products in an algal biorefinery approach, which is more economic than the single-product approach.
11.5.2
Pigments
Microalgae have a variety of pigments such as chlorophyll, lutein, carotenoids, and phycocyanin. Carotenoids from microalgae have a 40-carbon backbone that contains a bulky conjugated, double-bond system, which has a key role in photosynthesis and photo-protection [115]. Microalgal pigments are widely used as feed additives in aquaculture and poultry farming, and also as a natural colorant for pharmaceuticals, cosmetics, and food. Microalgal pigments are known for their antioxidant, antiinflammatory, antiobesity, antiangiogenic, anticancer, and neuroprotective activities [116]. b-Carotene, a vitamin A precursor, is used for its food coloring properties and antioxidant activities. It is known to counteract free radicals involved in gastrointestinal cancer, aging, and arthritis. b-Carotene has the largest share of the carotenoids market, valued at $285 million in 2015, with a compounded annual growth rate of 1.8% (www. oilgae.com, February 2015). Chemically synthesized b-carotene has stereogeometry of all-trans, which affects bioavailability. Dunaliella b-carotene is available in 9-cis and alltrans isomeric forms, which enhance bioavailability and beneficial properties. The first commercial algal pigment launched was Dunaliella salinaederived b-carotene and the successful outdoor culture was attributed to the ability of D. salina to grow in high salinity and accumulate high quantities of b-carotene [117]. Commercial plants in Australia (Aquacarotene and Betatene) use extensive, unmixed open ponds for the
340
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
Table 11.3 Industrial Cultivation of Microalgae for Production of Biofuels and Bio-Based chemicals Series Number Company 1.
Algaelink, The Netherlands
2.
Algasol, Bangladesh
3.
Algenol Biofuels, United States
4.
Aquaflow Bionomic, New Zealand
5.
Aquatic Energy, United States
6.
Aurora Algae, United States
7.
A2BE Carbon Capture, Colorado, United States Algae Tec. Ltd., Australia
8.
9. 10.
BFS Biofuel Systems, Spain Cellana, United States
Production Technology
Main Products
Horizontal and tubular photobioreactors based on polycarbonates with computerized process controls, technical support for algal biofuel companies, commercial supply of Nannochloropsis, Isochrysis, Tetraselmis, and Pavlova Proprietary floating photobioreactor made of thin, flexible, polymeric material to be deployed on land or in water. Proprietary flexible plastic film photobioreactor that facilitates product creation and collection Genetically engineered Synechocystis or Anabaena with heterologous ethanol-producing genes (Patent Application No. 20100297736) Open ponds Large-scale wild algae harvesting from open ponds Raceway ponds Scenedesmus obliquus (Patent Application No. 20110138682) Open ponds with a proprietary pond information management system Nannochloropsis for polyunsaturated fatty acid production (A2EPAPure fact sheet) Patented photobioreactor, two-bag system with rollers for mixing (US Patent US 20070048848 A1) Enclosed, modular, engineered bioreactor technology for industrial-scale cultivation of nongovernmental organization algae Vertical tubular photobioreactor with industrial CO2 capture Open ponds and tubular systems with turbulent flow regime Lyngbya lagerheimii, Spirulina platensis, Haematococcus pluvialis, Chlorella sorokiniensis, Nanofrustulum, and Tetraselmis (US Patent No. 5,541,056) Patented hybrid algae production system, basically covered pond system
Biomass, Jet fuel in the pipeline
11.
Green star, United Kingdom
12.
Heliae, United States Spiral photobioreactor with new oval glass tubes as building material H. pluvialis for astaxanthin
Biomass
Bioethanol
Biodiesel
Lipids for biodiesel, omega 3 oils Omega 3 oils, fuel, feed and protein
Biofuel
Biomass for biofuels
Biopetroleum Biofuels, Nutraceuticals, pharmaceuticals, aquaculture and animal feeds, and Omega-3 oils, Biodiesel, lubricants and engine additive, algae growth enhancer Astaxanthin, Phycoterra (natural crop enhancer)
Chapter 11 � Photobioreactors 341
Table 11.3 Industrial Cultivation of Microalgae for Production of Biofuels and Bio-Based chemicalsdcont’d Series Number Company 13. 14. 15. 16.
LiveFuels, United States Muradel, Australia Pond Biofuels, Canada Sapphire Energy, United States
17.
Seambiotic, Israel
18.
Solazyme, United States
19.
Solix Biosystems, United States
20.
Subitec, Germany
Production Technology
Main Products
Open ponds in estuarine environments, at intersection of rivers Open-paddlewheel mixed ponds Saline-tolerant, highelipid yielding microalgae Unique photobioreactor that uses exhaust gases as CO2 source Open pond systems Various genetically modified algal species for hydrocarbon, lipid production, and engineered saline-tolerant microalgae Chlamydomonas reinhardtii (US Patent No. 20090087890), Dunaliella salina, H. pluvalis, Scenedesmus dimorphus, Dunaliella viridis, and Dunaliella tertiolecta (http://stks.freshpatents.com/ Sapphire-Energy-Inc-nm1.php) Open pond systems and raceway ponds, with industrial gas as CO2 source Skeletonema, Nannochloropsis, Chlorococcum, Dunaliella, Nannochloris, and Tetraselmis (US Patent No. 20080220486 A1) Heterotrophic cultivation of algae in conventional bioreactors with agriculture-based sugars as feed Transgenic Prototheca (Patent Application No. 20100151538) Botryococcus braunii (Patent Application No. 20130071909) Proprietary algae growth system that incorporates Lumian photobioreactor panels (Diffuse light-extended surface area watersupported photobioreactor, US Patent No. 20080160591 A1) Nannochloropsis Proprietary flat-panel air-lift reactor with patented static mixers
Biodiesel Green crude, algae-based biofuels Biofuels, Biodiesel Biofuels, omega oils, highvalue proteins
Biodiesel
Jet fuel, renewable diesel, biodiesel, high-value algal oils and proteins
Biodiesel, green diesel, jet fuel, and bioplastics, omega oils and personal care products
Food, feed, fuel, pharmaceuticals, nutraceuticals
Data were retrieved from the corresponding Web sites of the company and their patent files as on October 2015.
cultivation of D. salina, and other firms in India, the United States, and China (Cyanotech, EID Parry, and Tainjin Lantai Biotechnology) use paddlewheel mixed raceway-type ponds for cultivation [118]. Astaxanthin, a powerful antioxidant known to reduce hyperpigmentation in skin, is used as a skin-lightening agent. The market value for astaxanthin is currently estimated at $200 million and is predicted to hit $700 million by 2017. It is commercially produced successfully from H. pluvialis, a freshwater unicellular green alga which can accumulate
342
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
astaxanthin up to 4% by weight and has the highest astaxanthin content from a natural source. The alga has a complex life cycle, with a vegetative green motile phase and a nonmotile cyst phase with a heavy cellulosic cell wall and high amounts of astaxanthin. Carotenogenic or astaxanthin accumulation can be induced by various stress conditions such as nutrient depletion or intense light. The algae can be grown in two stages: a green phase for biomass accumulation, always in a closed PBR to prevent contamination; and a second stress phase to induce astaxanthin accumulation, either in outdoor ponds (Cyanotech, Hawaii) or in closed systems (Alga Technologies, Israel). Lutein is a human serum carotenoid which is not synthesized by humans and thus must be obtained by ingesting lutein-rich food. Lutein is present in different forms in fruits, vegetables, and algae as all-trans-lutein, cis-lutein, epoxy-lutein, and lutein linked to proteins. It has been reported that the intake of lutein is important in preventing diseases such as some cancers [119,120], cardiovascular diseases [121], and degenerative human diseases [122,123]. The antioxidant effect of lutein may have a role in preventing age-related macular degeneration and other eye diseases. Microalgae that are known to produce lutein include Murellopsis sp., S. almeriensis, and Chlorella protothecoides (4.5e5.5 mg/g dry weight) [124]. Moreover, fucoxanthins, which are derived from brown algae, have the ability to counteract the oxidative stress induced by UV radiation and are used in cosmetic preparations [100].
11.5.3
Feed
Current supplies of animal proteins are inadequate to meet the predicted “protein gap” owing to rapidly depleting resources such as arable land and clean water [125]. The trend of increasing consumption of protein and decreasing natural harvests will dictate the derivation of a larger portion of seafood from aquaculture in the future. The ruminant sector will also need to increase production by fully using the available biomass from foraging, crop residues, agro-industrial by-products and other unconventional feed resources such as microalgae, yeasts, fungi, and bacteria. It has been recognized that phototrophic microorganisms (e.g., microalgae and cyanobacteria) can be used as a good protein source. Using microalgae as feed is promising because microalgae have a rapid growth rate, possess the highest photosynthetic capability per unit volume, do not require arable land, and can use brackish water and wastewater for growth. A comprehensive analysis of the gross chemical compositions of different microalgae is shown in Table 11.4. It clearly demonstrates that microalgae contain useful components and are thus full of potential for a variety of applications. However, to date, few microalgae have been produced successfully on a large scale. Successful examples mainly belong to the chlorophyceae Chlorella sp. and S. obliquus as well as the cyanobacteria Spirulina sp. and Athrospira sp. Chlorella is a spherical, eukaryotic, unicellular alga with a diameter of 5e10 mm. Its hemicellulosic cell wall accounts for the rigidity of the cells. The green alga Scenedesmus is similar in size but forms four-celled colonies. Spirulina sp. and Arthrospira sp. are photosynthetic,
Chapter 11 � Photobioreactors 343
Table 11.4 General Composition of Different Algae (Percent Dry Weight) [126] Microalgae
Protein
Carbohydrates
Lipids
Aphanizomenon flos-aquae Chlamydomonas rheinhardii Chlorella pyrenoidosa Chlorella vulgaris Dunaliella salina Porphyridium cruentum Scenedesmus obliquus Arthrospira maxima Spirulina platensis Synechococcus sp.
62 48 57 51e58 57 28e39 50e56 60e71 46e63 63
23 17 26 12e17 32 40e57 10e17 13e16 8e14 15
3 21 2 14e22 6 9e14 12e14 6e7 4e9 11
filamentous, spiral-shaped, multicellular organisms measuring around 0.5 mm long. They were formerly classified as blue-green algae but were later changed to cyanobacteria owing to their prokaryotic structure [126]. Spirulina is marketed as a powder or tablet and is produced commercially in open raceway ponds with a productivity of about 12 metric tons/ha/year in the United States, China, Japan, Taiwan, Thailand, and India [127]. The major hurdle of using microalgae as aquacultural feed is the difficulty in supplying microalgae as the primary food source for larval stages in some of the most valuable farmed fish and shellfish. Using live feed such as brine shrimp (Artemia) or rotifer (Brachionus) as a nutrition transporter is a traditional way to avoid the problems of ingestion and digestion in larvae of fish and shellfish. Alternatively, a feeding method that periodically adds small amounts of feed, called a microdiet, can be used to overcome the issue of overfeeding. However, this method is still a work in progress [128]. Microalgae appear to be nutritionally suitable as a resource for marine culture feed because many microalgae species have a high protein and PUFA content. Under suitable culture conditions, some microalgal species are able to accumulate 28e71% protein per dry weight (Table 11.4) [129]. Nutritional studies have demonstrated that algal proteins are of high quality compared with vegetable proteins [126].
11.5.4
Polyunsaturated Fatty Acids
Essential fatty acids are a group of fatty acids which cannot be synthesized by animals and humans because of a lack of necessary enzymes; they need to be supplemented in the diet. The traditional source of these fatty acids is fish oils obtained from oily fish such as salmon [117]. However, concerns have been raised about the accumulation of mercury and other toxic products by fish, which has restricted their consumption. Omega-3 fatty acids, that have a cis double bond at position 3, have received increasing attention owing to numerous health benefits. DHA, an omega-3 fatty acid, is essential for brain function and is important for development of the nervous system and visual abilities
344
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
during early development and infancy [127]. DHA is a widely used supplement in maternal health supplements and infant formulas. Martek Bioscience Corporation produces DHA commercially from Crypthecodinium cohini for infant formula. Other commercial producers of PUFA and the microalga used are listed in Table 11.3. EPA, arachidonic acid, and linolenic acid are some other PUFAs for which microalgal production will be available [130].
11.5.5
Cosmetic Products
Sunlight carries the fundamental energy for life on earth by driving photosynthesis, but UV radiation from the sun has clear detrimental effects. Solar radiation with a short wavelength has been shown to have ecologically significant effects on terrestrial, freshwater, and marine ecosystems. Anthropogenically released atmospheric pollutants such as chlorofluorocarbons are responsible for continued depletion of the stratospheric ozone layer, and consequently cause an increase in solar UV radiation. UV radiation is highly genotoxic but does not penetrate the body deeper than the skin. The sun emits UV radiation across a broad spectrum from the high-energy UVC band (wavelengths below 280 nm) and the UVB band (280e315 nm) to the UVA band (315e400 nm, bordering the visible band from 400 to 800 nm) [131]. Microalgae-based cosmetics are mainly based on the antioxidant activity of pigments and the nourishing effect of nutrients present in extracts such as proteins and lipids. They protect skin from the harmful effects of UV radiation and counteract oxidative damage. Polysaccharides derived from macroalgae are known for antiaging properties [100]. Many algae-based products are used as topical agents such as sunscreens; moisturizing agents; toners; and skin tightening, skin brightening and antiaging agents. Any regular visitor to a spa would readily recognize Spirulina packs used for skin lightening and wrinkle reduction. Solazyme, the algae company, launched the Algenist line of high-end cosmetics in 2011, based on Alguronic oil extracted from microalgae. Important microalgae used in cosmeceutical preparations include Spirulina sp., Chlorella sp., and Nannochloropsis sp., which are known for their high protein content and antioxidant activities. Mycosporine-like amino acids (MAAs) have been identified in taxonomically diverse organisms, including a marine heterotrophic bacterium, cyanobacteria, microalgae, and macroalgae. MAAs are one of nature’s sunscreens, with 21 structurally distinct MAAs currently identified in marine and terrestrial organisms [132]. Whereas their main role is in screening against positive UV radiation, MAAs also protect against sunlight damage by acting as antioxidant molecules, scavenging toxic oxygen radicals. Further roles include acting as compatible solutes to protect cells against salt stress, where they are involved in protecting against desiccation or thermal stress in certain organisms and as intracellular nitrogen reserves [133]. From a more industrially applied biotechnological perspective, MAAs are becoming important as compounds of interest to supplement and/or replace commercially available sunscreens, particularly because the requirement for natural
Chapter 11 � Photobioreactors 345
products and replacement of petrochemical-based products is becoming more stringent. From an applied perspective, the broadband absorption of the MAAs, particularly in UVA, and their high-UV absorption coefficients are attractive properties in the search for synthetic replacements [133].
11.6 Concluding Remarks The types and operating factors of PBRs used for the cultivation of microalgae to produce biofuels and chemicals with the additional benefits of CO2 reduction are described in this chapter. Because of the requirements of light energy and CO2 gas as the inorganic carbon source, the performance of microalgae growth and follow-up target product production greatly depend on the PBRs used for their cultivation. The main processes involved in commercial cultivation of microalgae for fuel or chemical production include strain selection, large-scale high-yield cultivation, extraction of the functional materials, and conversion to final products. Among them, the design of highly efficient large-scale cultivation systems (or PBRs) is of pivotal importance. Large-scale cultivation of microalgae forms the foundation of biofuel industries; many are resorting to open pond or raceway pond systems because of the high cost involved in installing and maintaining closed-type PBRs and specially designed PBRs. Thus, more efforts are still needed in developing more versatile, economic, and effective outdoor PBRs for large-scale cultivation of microalgae. The emerging algae-based biofuels and chemicals industry is proof of success of such studies and is expected to grow in the near future with the aid of advanced PBR technologies.
References [1] W. Martin, T. Rujan, E. Richly, A. Hansen, S. Cornelsen, T. Lins, D. Leister, B. Stoebe, M. Hasegawa, D. Penny, Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus, Proceedings of the National Academy of Sciences 99 (2002) 12246e12251. [2] H. Li, Z. Liu, Y. Zhang, B. Li, H. Lu, N. Duan, M. Liu, Z. Zhu, B. Si, Conversion efficiency and oil quality of low-lipid high-protein and high-lipid low-protein microalgae via hydrothermal liquefaction, Bioresource Technology 154 (2014) 322e329. [3] S.N. Naik, V.V. Goud, P.K. Rout, A.K. Dalai, Production of first and second generation biofuels: a comprehensive review, Renewable and Sustainable Energy Reviews 14 (2010) 578e597. [4] L. Brennan, P. Owende, Biofuels from microalgaeda review of technologies for production, processing, and extractions of biofuels and co-products, Renewable and Sustainable Energy Reviews 14 (2010) 557e577. [5] D.-T. Tran, C.-L. Chen, J.-S. Chang, Effect of solvents and oil content on direct transesterification of wet oil-bearing microalgal biomass of Chlorella vulgaris ESP-31 for biodiesel synthesis using immobilized lipase as the biocatalyst, Bioresource Technology 135 (2013) 213e221. [6] S.-H. Ho, P.-J. Li, C.-C. Liu, J.-S. Chang, Bioprocess development on microalgae-based CO2 fixation and bioethanol production using Scenedesmus obliquus CNW-N, Bioresource Technology 145 (2013) 142e149.
346
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
[7] C.-H. Liu, C.-Y. Chang, Q. Liao, X. Zhu, C.-F. Liao, J.-S. Chang, Biohydrogen production by a novel integration of dark fermentation and mixotrophic microalgae cultivation, International Journal of Hydrogen Energy 38 (2013) 15807e15814. [8] D. Beneroso, J.M. Bermu´dez, A. Arenillas, J.A. Mene´ndez, Microwave pyrolysis of microalgae for high syngas production, Bioresource Technology 144 (2013) 240e246. [9] H.-H. Cheng, L.-M. Whang, K.-C. Chan, M.-C. Chung, S.-H. Wu, C.-P. Liu, S.-Y. Tien, S.-Y. Chen, J.-S. Chang, W.-J. Lee, Biological butanol production from microalgae-based biodiesel residues by Clostridium acetobutylicum, Bioresource Technology 184 (2015) 379e385. [10] J.C.-W. Lan, K. Raman, C.-M. Huang, C.-M. Chang, The impact of monochromatic blue and red LED light upon performance of photo microbial fuel cells (PMFCs) using Chlamydomonas reinhardtii transformation F5 as biocatalyst, Biochemical Engineering Journal 78 (2013) 39e43. [11] T.M. Mata, A.A. Martins, N.S. Caetano, Microalgae for biodiesel production and other applications: a review, Renewable and Sustainable Energy Reviews 14 (2010) 217e232. [12] A.P. Carvalho, L.A. Meireles, F.X. Malcata, Microalgal reactors: a review of enclosed system designs and performances, Biotechnology Progress 22 (2006) 1490e1506. [13] A. Richmond, Z. Cheng-Wu, Optimization of a flat plate glass reactor for mass production of Nannochloropsis sp. outdoors, Journal of Biotechnology 85 (2001) 259e269. [14] T. Chrismadha, M.A. Borowitzka, Effect of cell density and irradiance on growth, proximate composition and eicosapentaenoic acid production of Phaeodactylum tricornutum grown in a tubular photobioreactor, Journal of Applied Phycology 6 (1994) 67e74. [15] A.S. Miron, A.C. Gomez, F.G. Camacho, E.M. Grima, Y. Chisti, Comparative evaluation of compact photobioreactors for large-scale monoculture of microalgae, Journal of Biotechnology 70 (1999) 249e270. [16] S.-H. Ho, C.-Y. Chen, D.-J. Lee, J.-S. Chang, Perspectives on microalgal CO2-emission mitigation systemsda review, Biotechnology Advances 29 (2011) 189e198. [17] S.A. Razzak, M.M. Hossain, R.A. Lucky, A.S. Bassi, H. de Lasa, Integrated CO2 capture, wastewater treatment and biofuel production by microalgae culturingda review, Renewable and Sustainable Energy Reviews 27 (2013) 622e653. [18] T.M. Iverson, Evolution and unique bioenergetic mechanisms in oxygenic photosynthesis, Current Opinion in Chemical Biology 10 (2006) 91e100. [19] S.-H. Ho, C.-Y. Chen, J.-S. Chang, Effect of light intensity and nitrogen starvation on CO2 fixation and lipid/carbohydrate production of an indigenous microalga Scenedesmus obliquus CNW-N, Bioresource Technology 113 (2012) 244e252. [20] Y. Chisti, Biodiesel from microalgae, Biotechnology Advances 25 (2007) 294e306. [21] X. Sun, Y. Cao, H. Xu, Y. Liu, J. Sun, D. Qiao, Y. Cao, Effect of nitrogen-starvation, light intensity and iron on triacylglyceride/carbohydrate production and fatty acid profile of Neochloris oleoabundans HK-129 by a two-stage process, Bioresource Technology 155 (2014) 204e212. [22] L. Christenson, R. Sims, Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts, Biotechnology Advances 29 (2011) 686e702. [23] K. Kumar, C.N. Dasgupta, B. Nayak, P. Lindblad, D. Das, Development of suitable photobioreactors for CO2 sequestration addressing global warming using green algae and cyanobacteria, Bioresource Technology 102 (2011) 4945e4953. [24] C. Posten, Design principles of photo-bioreactors for cultivation of microalgae, Engineering in Life Sciences 9 (2009) 165e177.
Chapter 11 � Photobioreactors 347
[25] P.H. Ravelonandro, D.H. Ratianarivo, C. Joannis-Cassan, A. Isambert, M. Raherimandimby, Influence of light quality and intensity in the cultivation of Spirulina platensis from Toliara (Madagascar) in a closed system, Journal of Chemical Technology and Biotechnology 83 (2008) 842e848. [26] S.-H. Ho, A. Nakanishi, X. Ye, J.-S. Chang, C.-Y. Chen, T. Hasunuma, A. Kondo, Dynamic metabolic profiling of the marine microalga Chlamydomonas sp. JSC4 and enhancing its oil production by optimizing light intensity, Biotechnology for Biofuels 8 (2015) 48. [27] Z.I. Khalil, M.M. Asker, S. El-Sayed, I.A. Kobbia, Effect of pH on growth and biochemical responses of Dunaliella bardawil and Chlorella ellipsoidea, World Journal of Microbiology and Biotechnology 26 (2010) 1225e1231. [28] A. Juneja, R.M. Ceballos, G.S. Murthy, Effects of environmental factors and nutrient availability on the biochemical composition of algae for biofuels production: a review, Energies 6 (2013) 4607e4638. [29] J.R. Coleman, B. Colman, Inorganic carbon accumulation and photosynthesis in a Bluegreen alga as a function of external pH, Plant Physiology 67 (1981) 917e921. [30] J.C. Goldman, M.R. Dennett, C.B. Riley, Effect of nitrogen-mediated changes in alkalinity on pH control and CO2 supply in intensive microalgal cultures, Biotechnology and Bioengineering 24 (1982) 619e631. [31] L. Rodolfi, G. Chini Zittelli, N. Bassi, G. Padovani, N. Biondi, G. Bonini, M.R. Tredici, Microalgae for oil: strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor, Biotechnology and Bioengineering 102 (2009) 100e112. [32] P.J. Hansen, Effect of high pH on the growth and survival of marine phytoplankton: implications for species succession, Aquatic Microbial Ecology 28 (2002) 279e288. [33] G. Breuer, P.P. Lamers, D.E. Martens, R.B. Draaisma, R.H. Wijffels, Effect of light intensity, pH, and temperature on triacylglycerol (TAG) accumulation induced by nitrogen starvation in Scenedesmus obliquus, Bioresource Technology 143 (2013) 1e9. [34] H.-W. Yen, S.-H. Ho, C.-Y. Chen, J.-S. Chang, CO2, NOx and SOx removal from flue gas via microalgae cultivation: a critical review, Biotechnology Journal 10 (2015) 829e839. [35] P. Westerhoff, Q. Hu, M. Esparza-Soto, W. Vermaas, Growth parameters of microalgae tolerant to high levels of carbon dioxide in batch and continuous-flow photobioreactors, Environmental Technology 31 (2010) 523e532. [36] K. Maeda, M. Owada, N. Kimura, K. Omata, I. Karube, CO2 fixation from the flue-gas on coal-fired thermal power-plant by microalgae, Energy Conversion and Management 36 (1995) 717e720. [37] S.-H. Ho, M.-C. Chan, C.-C. Liu, C.-Y. Chen, W.-L. Lee, D.-J. Lee, J.-S. Chang, Enhancing lutein productivity of an indigenous microalga Scenedesmus obliquus FSP-3 using light-related strategies, Bioresource Technology 152 (2014) 275e282. [38] D.S. Pisal, S.S. Lele, Carotenoid production from microalga, Dunaliella salina, Indian Journal of Biotechnology 4 (2005) 476e483. [39] A.E. Tjahjono, Y. Hayama, T. Kakizono, Y. Terada, N. Nishio, S. Nagai, Hyper-accumulation of astaxanthin in a green alga Haematococcus pluvialis at elevated temperatures, Biotechnology Letters 16 (1994) 133e138. [40] G.O. James, C.H. Hocart, W. Hillier, G.D. Price, M.A. Djordjevic, Temperature modulation of fatty acid profiles for biofuel production in nitrogen deprived Chlamydomonas reinhardtii, Bioresource Technology 127 (2013) 441e447. [41] L. Xin, H. Hong-Ying, Z. Yu-Ping, Growth and lipid accumulation properties of a freshwater microalga Scenedesmus sp. under different cultivation temperature, Bioresource Technology 102 (2011) 3098e3102.
348
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
[42] I.S. Suh, C.G. Lee, Photobioreactor engineering: design and performance, Biotechnology and Bioprocess Engineering 8 (2003) 313e321. [43] S.M. Renaud, L.V. Thinh, G. Lambrinidis, D.L. Parry, Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures, Aquaculture 211 (2002) 195e214. [44] C.-Y. Kao, T.-Y. Chen, Y.-B. Chang, T.-W. Chiu, H.-Y. Lin, C.-D. Chen, J.-S. Chang, C.-S. Lin, Utilization of carbon dioxide in industrial flue gases for the cultivation of microalga Chlorella sp., Bioresource Technology 166 (2014) 485e493. [45] E. Ono, J.L. Cuello, Carbon dioxide mitigation using thermophilic cyanobacteria, Biosystems Engineering 96 (2007) 129e134. [46] M. Calvin, 40 years of photosynthesis and related activities, Photosynthesis Research 21 (1989) 3e16. [47] X. Wang, Z. Ruan, P. Sheridan, D. Boileau, Y. Liu, W. Liao, Two-stage photoautotrophic cultivation to improve carbohydrate production in Chlamydomonas reinhardtii, Biomass and Bioenergy 74 (2015) 280e287. [48] J. Raven, Inorganic carbon acquisition by eukaryotic algae: four current questions, Photosynthesis Research 106 (2010) 123e134. [49] B. Wang, Y.Q. Li, N. Wu, C.Q. Lan, CO2 bio-mitigation using microalgae, Applied Microbiology and Biotechnology 79 (2008) 707e718. [50] S. Van Den Hende, H. Vervaeren, N. Boon, Flue gas compounds and microalgae: (Bio-) chemical interactions leading to biotechnological opportunities, Biotechnology Advances 30 (2012) 1405e1424. [51] E. Jacob-Lopes, C.H. Gimenes Scoparo, M.I. Queiroz, T.T. Franco, Biotransformations of carbon dioxide in photobioreactors, Energy Conversion and Management 51 (2010) 894e900. [52] P. Carlozzi, Closed photobioreactor assessments to grow, intensively, light dependent microorganisms: a twenty-year Italian outdoor investigation, The Open Biotechnology Journal 2 (2008) 63e72. [53] S.I. Blackburn, J.K. Volkman, Microalgae: A Renewable Source of Bioproducts, Wiley-Blackwell, New York, NY, USA, 2012. [54] G. Huang, F. Chen, D. Wei, X. Zhang, G. Chen, Biodiesel production by microalgal biotechnology, Applied Energy 87 (2010) 38e46. [55] F.A. Ferna´ndez, J.F. Sevilla, E.M. Grima, Photobioreactors for the production of microalgae, Reviews in Environmental Science and Bio/Technology 12 (2013) 131e151. [56] C.N. Dasgupta, J.J. Gilbert, P. Lindblad, T. Heidorn, S.A. Borgvang, K. Skjanes, D. Das, Recent trends on the development of photobiological processes and photobioreactors for the improvement of hydrogen production, International Journal of Hydrogen Energy 35 (2010) 10218e10238. [57] M.A. Borowitzka, N.R. Moheimani, Open Pond Culture Systems. Algae for Biofuels and Energy, Springer, 2013. [58] Y.-K. Lee, Microalgal mass culture systems and methods: their limitation and potential, Journal of Applied Phycology 13 (2001) 307e315. [59] R. Singh, S. Sharma, Development of suitable photobioreactor for algae productionda review, Renewable and Sustainable Energy Reviews 16 (2012) 2347e2353. [60] S.C. James, V. Boriah, Modeling algae growth in an open-channel raceway, Journal of Computational Biology 17 (2010) 895e906.
Chapter 11 � Photobioreactors 349
[61] O. Jorquera, A. Kiperstok, E.A. Sales, M. Embiruc¸u, M.L. Ghirardi, Comparative energy life-cycle analyses of microalgal biomass production in open ponds and photobioreactors, Bioresource Technology 101 (2010) 1406e1413. [62] C.U. Ugwu, H. Aoyagi, Microalgal culture systems: an insight into their designs, operation and applications, Biotechnology 11 (2012) 127. [63] J. Park, R. Craggs, A. Shilton, Wastewater treatment high rate algal ponds for biofuel production, Bioresource Technology 102 (2011) 35e42. [64] O. Pulz, Photobioreactors: production systems for phototrophic microorganisms, Applied Microbiology and Biotechnology 57 (2001) 287e293. [65] T. Kondo, T. Wakayama, J. Miyake, Efficient hydrogen production using a multi-layered photobioreactor and a photosynthetic bacterium mutant with reduced pigment, International Journal of Hydrogen Energy 31 (2006) 1522e1526. [66] C.-Y. Chen, G.D. Saratale, C.-M. Lee, P.-C. Chen, J.-S. Chang, Phototrophic hydrogen production in photobioreactors coupled with solar-energy-excited optical fibers, International Journal of Hydrogen Energy 33 (2008) 6886e6895. [67] E. Nakada, Y. Asada, T. Arai, J. Miyake, Light penetration into cell suspensions of photosynthetic bacteria and relation to hydrogen production, Journal of Fermentation and Bioengineering 80 (1995) 53e57. [68] S.H. Lee, B.H. Jo, S.Y. Shin, S.H. Lee, Higher biomass productivity of microalgae in an attached growth system, using wastewater, Journal of Microbiology and Biotechnology 24 (2014) 1566e1573. [69] T. Liu, J. Wang, Q. Hu, P. Cheng, B. Ji, J. Liu, Y. Chen, W. Zhang, X. Chen, L. Chen, Attached cultivation technology of microalgae for efficient biomass feedstock production, Bioresource Technology 127 (2013) 216e222. [70] P. Cheng, B. Ji, L. Gao, W. Zhang, J. Wang, T. Liu, The growth, lipid and hydrocarbon production of Botryococcus braunii with attached cultivation, Bioresource Technology 138 (2013) 95e100. [71] B. Wang, C.Q. Lan, M. Horsman, Closed photobioreactors for production of microalgal biomasses, Biotechnology Advances 30 (2012) 904e912. [72] M.A. Borowitzka, Commercial production of microalgae: ponds, tanks, tubes and fermenters, Journal of Biotechnology 70 (1999) 313e321. [73] A.S. Miro´n, M.-C.C. Garcıa, F.G. Camacho, E.M. Grima, Y. Chisti, Growth and biochemical characterization of microalgal biomass produced in bubble column and airlift photobioreactors: studies in fed-batch culture, Enzyme and Microbial Technology 31 (2002) 1015e1023. [74] G. Dragone, B.D. Fernandes, A.A. Vicente, J.A. Teixeira, Third generation biofuels from microalgae, in: A. Me´ndez-Vilas (Ed.), Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology, Formatex Research Center, Badajoz, Spain, 2010. [75] A.S. Miro´n, F.G. Camacho, A.C. Go´mez, E.M. Grima, Y. Chisti, Bubble-column and airlift photobioreactors for algal culture, Bioengineering, Food, and Natural Products 46 (2000) 1872e1887. [76] L. Xu, P.J. Weathers, X.R. Xiong, C.Z. Liu, Microalgal bioreactors: challenges and opportunities, Engineering in Life Sciences 9 (2009) 178e189. [77] R. Halim, B. Gladman, M.K. Danquah, P.A. Webley, Oil extraction from microalgae for biodiesel production, Bioresource Technology 102 (2011) 178e185. [78] K. Kaewpintong, A. Shotipruk, S. Powtongsook, P. Pavasant, Photoautotrophic high-density cultivation of vegetative cells of Haematococcus pluvialis in airlift bioreactor, Bioresource Technology 98 (2007) 288e295.
350
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
[79] M.J. Barbosa, M. Janssen, N. Ham, J. Tramper, R.H. Wijffels, Microalgae cultivation in air-lift reactors: modeling biomass yield and growth rate as a function of mixing frequency, Biotechnology and Bioengineering 82 (2003) 170e179. [80] J. Degen, A. Uebele, A. Retze, U. Schmid-Staiger, W. Tro¨sch, A novel airlift photobioreactor with baffles for improved light utilization through the flashing light effect, Journal of Biotechnology 92 (2001) 89e94. [81] S. Monkonsit, S. Powtongsook, P. Pavasant, Comparison between airlift photobioreactor and bubble column for Skeletonema costatum cultivation, Engineering Journal 15 (2011) 53e64. [82] Y. Chisti, M. Moo-Young, Improve the performance of airlift reactors, Chemical Engineering Progress (June 1993) 38e45. [83] S. Oncel, F.V. Sukan, Comparison of two different pneumatically mixed column photobioreactors for the cultivation of Artrospira platensis (Spirulina platensis), Bioresource Technology 99 (2008) 4755e4760. [84] S. Krichnavaruk, S. Powtongsook, P. Pavasant, Enhanced productivity of Chaetoceros calcitrans in airlift photobioreactors, Bioresource Technology 98 (2007) 2123e2130. [85] N. Zou, A. Richmond, Effect of light-path length in outdoor flat plate reactors on output rate of cell mass and of EPA in Nannochloropsis sp, Journal of Biotechnology 70 (1999) 351e356. [86] K. Zhang, S. Miyachi, N. Kurano, Evaluation of a vertical flat-plate photobioreactor for outdoor biomass production and carbon dioxide bio-fixation: effects of reactor dimensions, irradiation and cell concentration on the biomass productivity and irradiation utilization efficiency, Applied Microbiology and Biotechnology 55 (2001) 428e433. [87] J.C. Ogbonna, T. Soejima, H. Tanaka, An integrated solar and artificial light system for internal illumination of photobioreactors, Journal of Biotechnology 70 (1999) 289e297. [88] M. Javanmardian, B. Palsson, High-density photoautotrophic algal cultures: design, construction, and operation of a novel photobioreactor system, Biotechnology and Bioengineering 38 (1991) 1182e1189. [89] A.M. Kunjapur, R.B. Eldridge, Photobioreactor design for commercial biofuel production from microalgae, Industrial and Engineering Chemistry Research 49 (2010) 3516e3526. [90] H.B. Chen, J.Y. Wu, C.F. Wang, C.C. Fu, C.J. Shieh, C.I. Chen, C.Y. Wang, Y.C. Liu, Modeling on chlorophyll a and phycocyanin production by Spirulina platensis under various light-emitting diodes, Biochemical Engineering Journal 53 (2010) 52e56. [91] M. Heining, A. Sutor, S. Stute, C. Lindenberger, R. Buchholz, Internal illumination of photobioreactors via wireless light emitters: a proof of concept, Journal of Applied Phycology 27 (2014) 59e66. [92] Y. Chisti, Biodiesel from microalgae beats bioethanol, Trends in Biotechnology 26 (2008) 126e131. [93] L.-J. Ren, X.-J. Ji, H. Huang, L. Qu, Y. Feng, Q.-Q. Tong, P.-K. Ouyang, Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Applied Microbiology and Biotechnology 87 (2010) 1649e1656. [94] C.-Y. Chen, Y.-C. Chen, H.-C. Huang, C.-C. Huang, W.-L. Lee, J.-S. Chang, Engineering strategies for enhancing the production of eicosapentaenoic acid (EPA) from an isolated microalga Nannochloropsis oceanica CY2, Bioresource Technology 147 (2013) 160e167. [95] C.-Y. Chen, P.-C. Kao, C.-J. Tsai, D.-J. Lee, J.-S. Chang, Engineering strategies for simultaneous enhancement of C-phycocyanin production and CO2 fixation with Spirulina platensis, Bioresource Technology 145 (2013) 307e312. [96] C.U. Ugwu, H. Aoyagi, H. Uchiyama, Photobioreactors for mass cultivation of algae, Bioresource Technology 99 (2008) 4021e4028.
Chapter 11 � Photobioreactors 351
[97] A. Demirbas, M. Fatih Demirbas, Importance of algae oil as a source of biodiesel, Energy Conversion and Management 52 (2011) 163e170. [98] R. Slade, A. Bauen, Micro-algae cultivation for biofuels: cost, energy balance, environmental impacts and future prospects, Biomass and Bioenergy 53 (2013) 29e38. [99] A. Muller-Feuga, R. Le Gue´des, A. Herve´, P. Durand, Comparison of artificial light photobioreactors and other production systems using Porphyridium cruentum, Journal of Applied Phycology 10 (1998) 83e90. [100] H.-M.D. Wang, C.-C. Chen, P. Huynh, J.-S. Chang, Exploring the potential of using algae in cosmetics, Bioresource Technology 184 (2015) 355e362. [101] C.-Y. Chen, C.-Y. Chang, Y.-C. Lo, S.-H. Ho, J.-S. Chang, Enhancing biohydrogen production from Chlorella vulgaris FSP-E under mixotrophic cultivation conditions, Energy Procedia 61 (2014) 870e873. [102] H.-W. Yen, I.C. Hu, C.-Y. Chen, S.-H. Ho, D.-J. Lee, J.-S. Chang, Microalgae-based biorefinery e from biofuels to natural products, Bioresource Technology 135 (2013) 166e174. [103] R.P. Utomo, Y.-R. Chang, D.-J. Lee, J.-S. Chang, Lutein recovery from Chlorella sp. ESP-6 with coagulants, Bioresource Technology 139 (2013) 176e180. [104] J.-M. Park, A. Kondo, J.-S. Chang, C. Perry Chou, P. Monsan, Biorefineries, Bioresource Technology 135 (2013) 1. [105] A. Laviano, S. Rianda, A. Molfino, F. Rossi Fanelli, Omega-3 fatty acids in cancer, Current Opinion in Clinical Nutrition and Metabolic Care 16 (2013) 156e161. � Mi�san, I.L. Milovanovi� [106] D.J. Kova� c, J.B. Simeunovi� c, O.B. Babi� c, A.C. c, Algae in food and feed, Food and Feed Research 40 (2013) 21e32. [107] W.Q. Fu, O. Guomundsson, G. Paglia, G. Herjolfsson, O.S. Andresson, B.O. Palsson, S. Brynjolfsson, Enhancement of carotenoid biosynthesis in the green microalga Dunaliella salina with lightemitting diodes and adaptive laboratory evolution, Applied Microbiology and Biotechnology 97 (2013) 2395e2403. [108] L. Pirastru, M. Darwish, F.L. Chu, F. Perreault, L. Sirois, L. Sleno, R. Popovic, Carotenoid production and change of photosynthetic functions in Scenedesmus sp. exposed to nitrogen limitation and acetate treatment, Journal of Applied Phycology 24 (2012) 117e124. [109] D. Mitra, J.H. van Leeuwen, B. Lamsal, Heterotrophic/mixotrophic cultivation of oleaginous Chlorella vulgaris on industrial co-products, Algal Research 1 (2012) 40e48. [110] C.-Y. Chen, K.-L. Yeh, H.-Y. Chang, J.-S. Chang, Strategies to improve oil/lipid production of microalgae in outdoor cultivation using vertical tubular-type photobioreactors, Energy Procedia 61 (2014) 2755e2758. [111] K.-L. Yeh, C.-Y. Chen, J.-S. Chang, pH-stat photoheterotrophic cultivation of indigenous Chlorella vulgaris ESP-31 for biomass and lipid production using acetic acid as the carbon source, Biochemical Engineering Journal 64 (2012) 1e7. [112] A. San Pedro, C.V. Gonza´lez-Lo´pez, F.G. Acie´n, E. Molina-Grima, Outdoor pilot production of Nannochloropsis gaditana: influence of culture parameters and lipid production rates in raceway ponds, Algal Research 8 (2015) 205e213. [113] S.K. Wang, Y.R. Hu, F. Wang, A.R. Stiles, C.Z. Liu, Scale-up cultivation of Chlorella ellipsoidea from indoor to outdoor in bubble column bioreactors, Bioresource Technology 156 (2014) 117e122. [114] J. Singh, S. Gu, Commercialization potential of microalgae for biofuels production, Renewable and Sustainable Energy Reviews 14 (2010) 2596e2610. [115] B. Demmig-Adams, W.W. Adams, The role of xanthophyll cycle carotenoids in the protection of photosynthesis, Trends in Plant Science 1 (1996) 21e26.
352
CURRENT DEVELOPMENTS IN BIOTECHNOLOGY AND BIOENGINEERING
[116] R. Pangestuti, S.W. Kim, Biological activities and health benefit effects of natural pigments derived from marine algae, Journal of Functional Foods 3 (2011) 255e266. [117] M.A. Borowitzka, High-value products from microalgaedtheir development and commercialization, Journal of Applied Phycology 25 (2013) 743e756. [118] J.A. Del Campo, M. Garcı´a-Gonza´lez, M.G. Guerrero, Outdoor cultivation of microalgae for carotenoid production: current state and perspectives, Applied Microbiology and Biotechnology 74 (2007) 1163e1174. [119] P. Astorg, Food carotenoids and cancer prevention: an overview of current research, Trends in Food Science and Technology 8 (1997) 406e413. [120] D. Heber, Q.Y. Lu, Overview of mechanisms of action of lycopene, Experimental Biology and Medicine 227 (2002) 920e923. [121] J.H. Dwyer, M. Navab, K.M. Dwyer, K. Hassan, P. Sun, A. Shircore, S. Hama-Levy, G. Hough, X. Wang, T. Drake, C.N. Merz, A.M. Fogelman, Oxygenated carotenoid lutein and progression of early atherosclerosis: the Los Angeles atherosclerosis study, Circulation 103 (2001) 2922e2927. [122] C.J. Chiu, A. Taylor, Nutritional antioxidants and age-related cataract and maculopathy, Experimental Eye Research 84 (2007) 229e245. [123] F. Granado, B. Olmedilla, I. Blanco, Nutritional and clinical relevance of lutein in human health, British Journal of Nutrition 90 (2003) 487e502. [124] J.M. Ferna´ndez-Sevilla, F.G. Acie´n Ferna´ndez, E. Molina Grima, Biotechnological production of lutein and its applications, Applied Microbiology and Biotechnology 86 (2010) 27e40. [125] C. Devendra, R.A. Leng, Feed resources for animals in Asia: issues, strategies for use, intensification and integration for increased productivity, Asian-Australasian Journal of Animal Science 24 (2011) 303e321. [126] E.W. Becker, Micro-algae as a source of protein, Biotechnology Advances 25 (2007) 207e210. [127] K. Hudek, L.C. Davis, J. Ibbini, L. Erickson, Commercial products from algae, in: Algal Biorefineries, Springer, 2014, pp. 275e295. [128] E. Gisbert, A. Skalli, I. Ferna´ndez, Y. Kotzamanis, J.L. Zambonino-Infante, R. Fabregat, Protein hydrolysates from yeast and pig blood as alternative raw materials in microdiets for gilthead sea bream (Sparus aurata) larvae, Aquaculture 338e341 (2012) 96e104. [129] K. Chojnacka, F.J. Marquez-Rocha, Kinetic and stoichiometric relationships of the energy and carbon metabolism in the culture of microalgae, Biotechnology 3 (2004) 21e34. [130] J.J. Milledge, Commercial application of microalgae other than as biofuels: a brief review, Reviews in Environmental Science and Bio/Technology 10 (2011) 31e41. [131] F.R. de Gruijl, Skin cancer and solar UV radiation, European Journal of Cancer 35 (1999) 2003e2009. [132] T. Rezanka, M. Temina, A.G. Tolstikov, V.M. Dembitsky, Natural microbial UV radiation filterse mycosporine-like amino acids, Folia Microbiologica (Praha) 49 (2004) 339e352. [133] C.A. Llewellyn, R.L. Airs, Distribution and abundance of MAAs in 33 species of microalgae across 13 classes, Marine Drugs 8 (2010) 1273e1291. [134] J.U. Grobbelaar, Factors governing algal growth in photobioreactors: the “open” versus “closed” debate, Journal of Applied Phycology 21 (2009) 489e492.