*Institute of Chemical Physics, USSR Academy of Sciences, 117334 Moscow, USSR. **Physics ... On the basis of the model âpolymer chain in an array of obstaclesâ the influence of the topology effects on the ... Introduction. One of the most ...
Physica 140A (1987) 506-520 North-Holland. Amsterdam
DYNAMICS OF A POLYMER CHAIN IN AN ARRAY OF OBSTACLES S.K. NECHAEV*,
A.N. SEMENOV**
*Institute of Chemical Physics, USSR Academy **Physics Department,
and M.K. KOLEVA**
of Sciences,
117334
Moscow State University, 119899
Moscow,
Moscow,
USSR
USSR
Received 4 March 1986
On the basis of the model “polymer chain in an array of obstacles” the influence of the topology effects on the dynamics of concentrated polymer systems is investigated theoretically. The 1 /z-expansion (where z is the coordinational number of the lattice of obstacles) is proposed for this problem. By means of this expansion the diffusion coefficient of a linear unclosed polymer chain is calculated. The equilibrium properties of linear closed chain (i.e. ring) unentangled with either of the edges of the lattice are investigated in detail. In particular, it is shown that the diffusion coefficient D of the center of mass of closed chain consisting of N links is proportional to N-5’2.
1. Introduction One of the most difficult problems in the elaboration of the microscopic theory of nonequilibrium properties of concentrated polymer systems is the description of topological interactions (or, in other words, restrictions on the motion of polymer chains, due to their mutual impenetrability)‘.*). The presence of physical entanglements of the chains leads to an essential change of dynamic macroscopic properties of polymer solutions and melts (for instance, the diffusion coefficient, the maximal relaxation time). Since the exact microscopic description of the entanglement effects is a very complicated problem, a model “polymer chain in a tube” has been proposed and studied in detai13’4). This model is now a generally accepted basis for the description of the dynamic properties of polymer solutions and meltssz6). At the same time it has become clear that the model “polymer chain in a tube” in its canonical form is imperfect in some respects and needs to be modified7-g,‘6). The alternative approach to the construction of the microscopic theory of the dynamics of strongly entangled polymer chains was suggested in the publications by Iwata with collaborators lo-12). Using the method of the Gauss linking 0378-4371 I87 /$03.50 0 Elsevier Science Publishers B .V (North-Holland Physics Publishing Division)
DYNAMICS OF A POLYMER CHAIN
507
number, generalised for dynamic problems, they had calculated the diffusion coefficient of the center of mass of a linear polymer chain in a solution. Unfortunately, rather complicated mathematical apparatus and adoption of some uncontrolled assumptions make this method unacceptable for the solution of many other problems of the dynamics of polymer systems. Moreover, at present there is no connection between Iwata's method and the model "polymer chain in a tube". We believe that one of the most useful and powerful models is the model "polymer chain in an array of obstacles"12-18), shown in fig. 1. The detailed description of this model as well as its superiority in comparison with the model "polymer chain in a tube" were presented in ref. 16. The representation of the polymer chain in an array of obstacles was used from the very beginning for the illustration of the model "polymer chain in a tube" but the direct investigation of this model is the topic of only few recent publicat i o n s - either analytical 13-~6) or devoted to computer experiments17-~9). Since this model is more detailed than the model "polymer chain in a tube" it is possible to use it for the detailed investigation of the influence of the topological restrictions on the motion of the chain. This investigation may be regarded as a further step in the construction of the microscopic theory of topological interactions. In this article within the framework of the model "polymer chain in an array of obstacles" we calculate diffusion coefficients of linear unclosed polymer chain and linear closed chain unentangled with either of the edges of the lattice. The detailed calculations of the partition function and of the radius of gyration
a /
/ P
/ AY
b
I
I
/
I
I/
I
'
I
]
x
I ] ."
I i
I
/r"i
Jl I
J
I~
~--!-j/
x
x
I
r-r--I I /t
__
x
x
' lJ
x
/ .
x
| .
.
~.
x
x
.
x
7
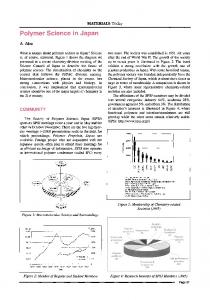
Fig. 1. Polymer chain in the lattice of obstacles: (a) three-dimensional case; (b) two-dimensional case.
508
S.K. N E C H A E V et al.
of the closed chain will be performed as well. Their knowledge is necessary for the investigation of the diffusion motion of a macromolecule. For the solution of these problems, the approach proposed in ref. 16 is applied.
2. Equilibrium characteristics of a polymer chain in the lattice of obstacles
Let us consider the linear polymer chain without volume interactions, placed into the lattice of obstacles, which are impenetrable for the chain. Let z be the coordination number of the lattice. We will consider here only the simplest case when the length of an elementary step of the chain, a is equal to the spacing of the lattice, c (see fig. 2). The exact consideration of the general case c ~ a is a somewhat more difficult problem and it will be presented in a separate publication. The condition c = a does not lead to the loss of generality of the obtained results, because (as shown in refs. 13 and 16) we can easily pass from the case c = a to the case c ~ a using simple scaling considerations. Let us fix the starting point (A) of the chain and the end of it (B). Now let us consider only those conformations that can be transformed into one another continuously (without crossing the edges of the lattice). These conformations belong to the class of topologically equivalent ones. It is possible to assign to any such class a point on the Cayley tree with the corresponding coordination number, z 16). Fig. 3 shows the walk on the Cayley tree corresponding to the walk in the real space (fig. 2). It should be noted that the analogy with the walk on the tree is valid for the case c ~ a as well. In this case the tree has a complex structure with a few branches connecting neighbouring nodes. The tree-like structure appears also in the continuous consideration (see ref. 16). If we eliminate all the double-folded sections of the chain we will obtain a sequence of steps which is called a primitive path of the chain (the dotted line in fig. 2). The concept of the primitive path is widely used both in studies of I
I
~
I
3t
xg
x
, L,
-
I I
_
I
,
.=,JI
t
y,
x
I
T
'__
I -r
y.
~
-
-'ri
~.
-
I
X
I
x
Fig. 2. Unclosed polymer chain in the lattice of obstacles: lattice realization in two-dimensional case, c = a. Primitive path is shown by dashed line.
DYNAMICS OF A POLYMER CHAIN
509
Fig. 3. Polymer macromolecule on the Cayley tree lattice.
the model "polymer chain in a tube ''5) and in the framework of the present modella'14A6). It can be easily shown that the consideration of the primitive path plays the role of the full topological invariant for the chain with fixed ends. In particular, the length of the primitive path, k, is an important topological invariant. Now we will calculate the distribution function for this value. Let PN(k) be the probability that the length of the primitive path of the random walk consisting of N steps in equal to k. Using the representation of the polymer chain on the Cayley tree it is easy to note that the value of PN(k) determines the probability of the fact that the unclosed N-step walk will finish at a distance of k steps on the tree from the origin (i.e. the place where initial link of the chain is fixed). The exact recurrence relation for the function PN(k) has the form (boundary and initial conditions included)
Pu(k)= z - 1
pN_a(k_l)+ 1 PN_l(k+l )
Z
k~2
Z
PN(1 ) -~- P N _ I ( 0 ) q- 1 P N _ I ( 2 ) ) Z
PN(O) = 1 PN ~(1) - -
--
(2.1)
)
Z
Po( k ) = ~k,o " The system of equations (2.1) can be simplified using the substitution eN(k)
-~- z - N ( z
--
1)~/2+N/Z2NQN(k),
k ~ O, (2.2)
PN(O ) = z-N(z _ 1)U/2 Z -- 1 2NQN(O), Z
510
S.K. NECHAEV
et al.
which leads to the relations QN( k ) = ~1 Q u - , ( k - 1 ) + z
QN(O) = ½ ~
½QN_I(k + I)
QN(1),
(2.3)
z
Q ° ( k ) - z - 1 6k'° "
If we consider k as a coordinate then eqs. (2.3) describe the random walk of N steps on the semi-infinite line k > 0. The solution of eqs. (2.3) in the limit N>>I is z
2z Qu(k) =
2-NNI 2 - 2/'\
2 +
k + -z - 2 !
N-k
is integer,
z--L-2-2
N-k
0,
is not integer. (2.4)
2
Later on we will use the smoothed function, Q n ( k ) , equal to one-half of the function (2.4) at any value of k (irrespective of the value of ( N - k)/2). Substituting eq. (2.4) into eq. (2.2) we find the asymptotic form of the value PN(k) near the maximum of the distribution (compare with ref. 13)
z
1
PN(k) - 2V~-S-1 2V2~N exp
[
z2 8 N ( z - 1)
(
)2] k
z -z 2 N
.
(2.5)
Hence the average length of the primitive path of the unclosed N-link chain is equal to z-2 (k) = - -
N.
(2.6)
z
A primitive path can have a lot of configurations in space. The only condition being that any following step of the primitive path should not be exactly opposite to the previous one. Therefore the mean-square distance (R a) between the ends of the primitive path of k steps is equal to (see ref. 20) z
(R2) - z-2
ka2 "
(2.7)
DYNAMICS OF A POLYMER CHAIN
511
For the mean-square distance between the ends of the linear unclosed N-link chain eqs. ( 2 . 6 ) - ( 2 . 7 ) give the obvious result ( R 2) = N a 2. Now we will consider the closed macromolecule placed in the regular array of obstacles. The fact that the chain is unentangled with either of the obstacles means that the length of the primitive path equals to zero (fig. 4). Hence the full partition function of closed unentangled chain Z u is equal to Z N = zUPN(O) = (z - 1) N/2 z - 1 2UQN(0). 7.
(2.8)
Substituting eq. (2.4) into eq. (2.8) we obtain the result (see also ref. 13) ZN=
(2~)NN
z
(2.9)
-3/2 .
Eq. (2.9) can be c o m p a r e d with the partition function of the closed macromolecule in the three-dimensional space without any topological restrictions, ~7> 1. As follows from eq. (2.6) the average stored length is ff = N - ( k ) = 2 / z . N; thus in the limit z >> 1 a part of the links in the section of stored length is small: if~N= 2/z ~ 1. Consequently, the polymer
f
o-3 ,!
c) Fig. 5. Fragment of the primitive path of the polymer chain with an elementary kink of stored length on it. Figs. a, h, c correspond to different positions of the kink.
514
S.K. NECHAEV et al.
chain conformation can be represented as a bare primitive path with a small number of side branches. The main contribution to diffusion in this case is given by the simplest branch in the double-folded form (on fig. 2 they are marked by the numbers 1, 2, 4). The concentration of these structures is equal to p = 1/z. It can be easily shown that the concentration of the branches of type 3 in fig. 2 is equal to 1 / z 2. The concentration of more complicated structures is even smaller. Therefore, in the first order of I/z-expansion such structures can be neglected. For further considerations it will be necessary to know the diffusion coefficient F of the motion of the simplest branch (or the kink) along the primitive path of the chain. Let us suppose that p~ is the probability of the fact that a kink is found on the nth link of the primitive path and is placed parallel to this path (fig. 5b), and q~ is the probability of the unparallel conformation (fig. 5c). For the values of p~ and qn we can write the following equations: OPn 3t
_
To
( qn + qn-1 + P . + I + P . - 1 - 2 p . ( z - 1))
z
(n¢O,N)
(3.1)
Oq. _ To ((z - 2)(p,, + P,,+I) - 2q,,). 3t z
After the standard transformations eq. (3.1) can be reduced to the equation Oan - 1~ --02~'~n Ot
(~'~n - - P . + q.) ,
(3.2)
On z
where the coefficient F is calculated in the main order of 1/z-expansion 70 F= 2(z-l)
_ Y0 2z "
(3.3)
It should be noted that in eq. (3.3) we neglect the interactions between kinks, because these interactions are so weak that they practically do not perturb the ground state of the system considered (the state in which the interactions between kinks are neglected). The correction to the diffusion coefficient F due to the interaction between the kinks has the order of magnitude 0 ( 1 / z 2) and this value in eq. (3.3) can be neglected. The motion of the kinks along the primitive path is identical to the spreading of the spin wave in the one-dimensional Ising model. In this case the 1/z-expansion is identical to the low-temperature expansion (see, for example, ref. 22). Diffusion coefficient, D, of the center of mass of a polymer chain can be found using the Einstein relation D = Tff,
(3.4)
DYNAMICS OF A POLYMER CHAIN
515
where ~ is the mobility of the chain. If we put the macromolecule in the external field, U, then each segment of the chain will have the energy u = - f . r / N , where f is the external force acting on this segment and r is the position of the segment in space. In the case of weak forces, which do not perturb the shape of the polymer chain, the following relation is fulfilled (see, for example, ref. 23): (V) = ( f ,
(3.5)
where (V) is the average velocity of the motion of center of mass of the chain. It is obvious that the action of external field induces the flow of the kinks of stored length along the primitive path, J. The velocity of the displacement of the center of mass of the chain is described by the continuity relation (3.6)
V= 2JR~N,
where R is the end-to-end vector of the chain and the coefficient 2 in eq. (3.6) appears because the stored length segment consists of two links of the chain. The energy dissipation due to the flow of kinks along the whole primitive path, J, is equal to dE T(J_] dt = ~ \pl
2
2z z-2 = T -- J2z 3'0 z
p(k)
-
-
2Tj2N U = - 3'o
z(z -
2).
(3.7)
The ratio T / F plays the role of the friction coefficient and is the concentration of elementary kinks. The work of external field per unit time is equal to dA dt = ( f ' V ) "
(3.8)
Substituting eq. (3.6) into eq. (3.8) and comparing the result obtained with eq. (3.7) we get for the flow J the following relation: J = 3,0R . f / ( N T z ( z
2)).
-
(3.9)
Eq. (3.9) can be substituted into eq. (3.6). After averaging we obtain for the mobility = 2%aZ/(3NZTz(z
-
2)).
(3.10)
Thus, in the main order of 1/z-expansion for the diffusion coefficient D we obtain
516
S.K. NECHAEV et al. D-
27°a2 3N2z 2 .
(3.11)
Eq. (3.11) generalizes the well-known expression for the diffusion coefficient obtained within the framework of the model "polymer chain in a tube"5). As it can be seen from eq. (3.11) if z---~ oo the diffusion coefficient tends to zero. This fact is due to the mechanism of motion considered. Actually, if z--~ ~, the equilibrium length of the primitive path is equal to the full contour length of the chain and the concentration of the kinks tends to zero; in this case all motions of the chain are effectively blocked up. It is noteworthy that the proposed method is quite general: if we take into account the interactions between the kinks and interactions between the kinks and external surrounding properties such as viscoelasticity of polymer chains in array of obstacles can be investigated.
4. Diffusion of closed unentangled macromolecule Let us describe first the main qualitative ideas which will enable us to find the dependence of the diffusion coefficient D of the center of mass on the length of the closed chain which is unentangled with either of the edges of the lattice. Closed unentangled polymer chain (see fig. 4) visits any cell of the lattice an even number of times. A double-cut of the chain in one of the cells (for example the cut in the points A and B on fig. 4) in general divides the macromolecule into two unequal parts. Let N 1 and N 2 be the numbers of the links in the smaller and greater parts of the chain, respectively. It is clear that the macromolecule can abandon a given cell only if the links of the macromolecule overflow from one part to another. The maximal time of this process, Zmax, must have the same order as the time of a chain diffusion at a distance of the order of the spacial size of the chain ~'ma,, ~ R R / D .
(4.1)
The diffusion of the chain is due to the fluctuations of the parts of stored length. It is obvious that the diffusion process can be considered as quasistationary because we are interested only in the large-scale motions of the whole chain24). The "overflowing" time will be maximal if a polymer chain is divided into two equal parts. It can be shown that for any equilibrium conformation of the macromolecule a division with 1