The FASEB Journal article fj.12-216176. Published online October 15, 2012.
The FASEB Journal • Research Communication
Tryptophan within basic peptide sequences triggers glycosaminoglycan-dependent endocytosis Chérine Bechara,*,†,‡ Manjula Pallerla,*,†,‡ Yefim Zaltsman,*,†,‡ Fabienne Burlina,*,†,‡ Isabel D. Alves,§ Olivier Lequin,*,†,‡ and Sandrine Sagan*,†,‡,1 *Université Pierre et Marie Curie (UPMC), Université Paris 6, Unité Mixte de Recherche (UMR) 7203, †Centre National de la Recherche Scientifique (CNRS), UMR 7203, and ‡Ecole Normale Supérieure (ENS), UMR 7203, Laboratoire des BioMolécules (LBM), Paris, France; and §Chimie et Biologie Des Membranes et Des Nanoobjets (CBMN), UMR 5248, CNRS, Pessac, France
Cell-penetrating peptides (CPPs) are short basic peptides (8 –30 aa) that can enter cells. CPPs use a
combination of multiple mechanisms for cellular entry, covering both direct translocation across the plasma membrane and endocytic processes (1, 2). For direct translocation, it is postulated that CPPs interact directly with the lipid bilayer, whereas endocytic processes can involve other membrane partners, such as proteoglycans. However, the molecular basis that controls these internalization processes is still not fully elucidated. Although there are few examples of pure hydrophobic CPP sequences, basic amino acids in the peptide sequence are generally required for interaction with the cell surface and entry (3). Thus, as a prerequisite step for peptide entry, electrostatic interactions should take place between the positively charged peptides and negatively charged molecules at the cell surface. These electrostatic interactions involve, in particular, plasma membrane phospholipids for membrane translocation and glycoconjugates, such as sialic acids (SAs) and glycosaminoglycans (GAGs), for endocytosis. SAs are typically found at the terminal ends of carbohydrate chains on the cell surface. Because of their negative charge, SAs are involved in the binding of positively charged molecules that can be further transported into cells. SAs contribute in various biological and pathological phenomena, including intercellular adhesion, signaling, and microbial attachment (4, 5). For instance, removal of SAs present at the surface of neuronal cells strongly reduced the uptake of the homeodomain sequence (60 aa) of antennapedia protein (6). However, this was not the case for the homeodomain-derived cell-penetrating peptide penetratin, which instead was trapped by SAs at the cell surface (7). As for GAGs, they have important roles in endocytosis and cellular signaling. They are, for example, involved in the binding of proteins to their receptors, an interaction that leads to the internalization of the complexes
Abbreviations: CD, circular dichroism; CPP, cell-penetrating peptide; CS, chondroitin sulfate; DLS, dynamic light scattering; FCS, fetal calf serum; GAG, glycosaminoglycan; HI, heparin; HS, heparan sulfate; ITC, isothermal titration calorimetry; MALDI, matrix-assisted laser desorption/ionization; MS, mass spectrometry; SA, sialic acid; TOF, time of flight; Trp, tryptophan; WT, wild-type
1 Correspondence: Université Pierre et Marie Curie, Laboratoire des Biomolécules, UMR 7203 CNRS, ENS, Case courrier 182, 4 place Jussieu, 75252 Paris cedex 05, France. E-mail:
[email protected] doi: 10.1096/fj.12-216176 This article includes supplemental data. Please visit http:// www.fasebj.org to obtain this information.
Deciphering the structural requirements and mechanisms for internalization of cell-penetrating peptides (CPPs) is required to improve their delivery efficiency. Herein, a unique role of tryptophan (Trp) residues in the interaction and structuring of cationic CPP sequences with glycosaminoglycans (GAGs) has been characterized, in relation with cell internalization. Using isothermal titration calorimetry, circular dichroism, NMR, mass spectrometry, and phase-contrast microscopy, we compared the interaction of 7 basic CPPs with 5 classes of GAGs. We found that the affinity of CPPs for GAGs increases linearly with the number of Trp residues, from 30 nM for a penetratin analog with 1 Trp residue to 1.5 nM for a penetratin analog with 6 Trp residues for heparin (HI); peptides with Trp residues adopt a predominantly -strand structure in complex with HI and form large, stable -sheet aggregates with GAGs; and in the absence of any cytotoxicity effect, the quantity of peptide internalized into CHO cells increased 2 times with 1 Trp residue, 10 times with 2 Trp residues, and 20 times with 3 Trp residues, compared with ⴙ6 peptides with no Trp residues. Therefore, Trp residues represent molecular determinants in basic peptide sequences not only for direct membrane translocation but also for efficient endocytosis through GAGs.—Bechara, C., Pallerla, M., Zaltsman, Y., Burlina, F., Alves, I. D., Lequin, O., Sagan S. Tryptophan within basic peptide sequences triggers glycosaminoglycan-dependent endocytosis. FASEB J. 27, 000 – 000 (2013). www.fasebj.org
ABSTRACT
Key Words: aggregation 䡠  strand 䡠  sheet 䡠 ␣ helix 䡠 membrane translocation
0892-6638/13/0027-0001 © FASEB
1
TABLE 1. Sequence of the carboxamide peptides (biotin-Gly4 at the N terminus) used in this study, with the number of basic (R,K) and tryptophan (W) residues
Sequence
R,K (n)
W (n)
RRRRRRRRR YGRKKRRQRRR RRLLRRLRR RQIKIFFQNRRMKWKK RQIKIWFQNRRMKWKK RRWWRRWRR RRWWRRWRRWRRWWRR
⫹9 ⫹8 ⫹6 ⫹7 ⫹7 ⫹6 ⫹10
0 0 0 1 2 3 6
Peptide
R9 TAT(47–57) R6L3 (W48F)-penetratin Penetratin R6W3 R10W6
(8 –11). GAGs are also implicated in the internalization of CPPs through an endocytic process (12–15). The cell surface displays different kinds of GAGs that are attached to core proteins, including heparan sulfate (HS) and chondroitin sulfate (CS), but few studies have assessed the role of these different GAGs in the interaction with CPPs in vitro and in cell internalization. Interactions of TAT, penetratin, oligoarginine, and other well-studied CPPs with HS and the structurally related heparin (HI) mainly revealed tight binding, leading to the formation of aggregates thought to be the result of numerous HI chains noncovalently crossed-linked by CPPs (16). Clusters formed with penetratin are more stable than those formed with TAT peptide (17). Penetratin preferentially binds CS, and the removal of endogenous CS reduces the internalization of penetratin peptides (18), whereas there is a specific HS proteoglycan epitope necessary for endo-
cytic stimulation and to which TAT peptide is able to bind (19). In addition to cationic residues, tryptophan (Trp) has emerged as an important amino acid for membrane translocation through its interaction with the phospholipid bilayer, but its involvement in GAG binding and thus in endocytic pathways has not yet been considered (20 –23). In this study, we have investigated the interaction and structuring of different cell-penetrating peptides [TAT, R9, R6L3, R6W3, penetratin, a mutant (W48F)-penetratin analog, and R10W6; Table 1] with HI and monosulfated (CSA and CSC) and disulfated (CSD and CSE) CSs (Fig. 1). The results demonstrate that incorporation of Trp residues in basic peptide sequences is a way to control the efficiency of internalization of CPPs and that Trp residues are molecular determinants, which strengthen the interaction of basic peptide sequences with negatively charged GAGs and enhance the amount internalized by endocytosis.
MATERIALS AND METHODS
Materials Peptides [containing an N-terminal biotin-(Gly)4 moiety and carboxamide at the C terminus] were obtained from PolyPeptide Laboratories (Strasbourg, France) or synthesized using the Boc solid-phase strategy. Unfractionated sodic HI was bought from Sanofi-Aventis (Bridgewater, NJ, USA). CSs were purchased from Seikagaku (Tokyo, Japan): CSA from sturgeon notochord, CSC
Sulfur Average MW (kDa) Content (%)
Major disaccharide sequence
Negative charge/GAG -OOC O
CSA
37.5
6.3
150
OSO3- OH
O
O O
HO
O
OH HN O -OOC O
CSC
60
6.69
250
OH
O
OSO3 O
O
HO
O
OH HN O
Figure 1. Chemical structures of the glycosaminoglycans used in this study.
-OOC O
CSD
30
7.1
100
OH
O
OSO3 O
O
HO
O
OSO3HN O -OOC O
CSE
62.5
8.75
260
OSO3- OSO3-
O
O O
HO
O
OH HN O
-OOC
HI
13
11.3
100
O
O
O
HO OSO3-
2
Vol. 27
February 2013
The FASEB Journal 䡠 www.fasebj.org
OSO3-
O
HO
O NHSO3-
BECHARA ET AL.
and CSD from shark cartilage, and CSE from squid cartilage. Tetramethylrhodamine isothiocyanate (TRITC)-streptavidin and avidin were purchased from Zymed Laboratories (Invitrogen, Carlsbad, CA, USA). Phalloidin-FITC, SA, and all other reagents were from Sigma-Aldrich (St. Louis, MO, USA). Cell culture Wild-type (WT) CHO-K1 cells, xylose transferase- or GAGdeficient CHO-pgsA745 (GAGneg) cells, and SA-deficient CHO-lec2 (SAneg) cells were obtained from American Type Culture Collection (Rockville, MD, USA). All cell lines were grown in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum (FCS), penicillin (100 IU/ml), streptomycin (100 IU/ml), and amphotericin B (1 g/ml) in a humidified atmosphere containing 5% CO2 at 37°C. Measure of cellular uptake and quantification of membrane-bound peptide Cellular uptake was quantified by matrix-assisted laser desorption/ionization (MALDI)-time of flight (TOF) mass spectrometry (MS) as described previously (24, 25). In this protocol, the peptides bear a tag on the N terminus composed of 4 glycine residues together with a biotin moiety for purification purposes (1H-peptide). This enables us to obtain an internal standard for absolute quantification by MS, having the same sequence as the one to quantify, except that it bears 4 deuterated glycine residues (2H-peptide). Adherent and confluent cells (106 cells, seeded the day before in 12-well plates) were incubated with the cell-penetrating 1H-peptide (1, 3, 5, 7, and 10 M) in culture medium (without FCS) for 75 min at 4 or 37°C. After washings, trypsin (37°C) or pronase (4°C) was added to degrade the remaining extracellular and membrane-bound peptide and to detach cells (12). The cells were then lysed and boiled in a solution containing a controlled and relevant quantity of the 2H-peptide. The lysate was incubated with streptavidin-coated magnetic beads to extract the peptides. For membrane-bound peptide quantification, no protease was added, and the cells were directly lysed after washings. The peptides were eluted from the streptavidincoated magnetic beads with ␣-cyano-4-hydroxycinnamic acid matrix and spotted on the MALDI plate. The samples were analyzed by MALDI-TOF MS (positive ion reflector mode) on a Voyager-DE Pro mass spectrometer (Applied Biosystems, Foster City, CA, USA). For each experiment, we used duplicate or triplicate wells, and the experiments were repeated independently ⱖ3 times. Isothermal titration calorimetry (ITC) ITC experiments were performed on a nano-ITC calorimeter (TA Instruments, New Castle, DE, USA). Titrations were performed by injecting 10-l aliquots of polysaccharides (HI, CSA, CSC, CSD, and CSE) into the calorimeter cell containing the peptide solution, with 5-min injection intervals. Peptides and polysaccharide solutions were used at different concentrations according to the peptide sequence and the GAG species (varying between 50 and 200 M for the peptides, 10 and 30 M for CSs, and 70 and 120 M for HI). The experiments were performed at 37°C in 50 mM Tris-Cl, 2 mM EDTA, and 150 mM NaCl, pH 7.5 (26, 27). Control experiments were performed by injection of buffer into peptide solution and injection of GAGs into buffer. Data analysis was performed with NanoAnalyze software provided by TA Instruments. Experiments were done ⱖ3 times. CATIONIC CELL-PENETRATING PEPTIDE INTERNALIZATION
Circular dichroism (CD) The CD spectra of the peptides were recorded using a J-815 spectrophotometer (Jasco, Tokyo, Japan). The spectra were acquired in a 1-mm pathlength at 20°C in a quartz optical cell, from 190 to 270 nm with a 0.2-nm step. Five scans were accumulated and averaged after subtraction of the baseline for an appropriate blank sample. The spectra were acquired for peptide concentrations varying from 10 to 50 M (corresponding to peptide totally bound to HI or peptide in excess with respect to HI, according to the stoichiometry of binding obtained by ITC) in 10 mM sodium phosphate buffer (pH 7.4) or in the presence of HI. CD measurements were reported as mean residual ellipticity (; deg · cm2 · dmol⫺1 · n⫺1). Secondary structure content was estimated from CD spectra using the deconvolution program CDFriend (28). The experimental error is estimated to be ⫾5% (28). NMR Experiments were performed on a Avance III spectrometer equipped with a TCI 1H/13C/15N cryoprobe (Bruker, Newark, DE, USA) and operating at a 1H frequency of 500 MHz. Peptide samples were prepared at a concentration of 0.1 mM in 3-mm NMR tubes (Norell, Inc., Landisville, NJ, USA) containing 170 l of PBS at pH 5.5, 10% D2O, and 0.1 mM sodium 2,2-dimethyl-2-silapentane-5-sulfonate (Isotec, St. Louis, MO, USA) for chemical shift calibration; pH is classically decreased to 5.5 to observe the amide protons of peptides on NMR spectra. Peptide samples were titrated with increasing concentrations of HI tetrasaccharide DP4 (Dextra, Reading, UK) from a 1 mM stock solution. NaCl was added in the samples using a 5 M stock solution. One-dimensional NMR experiments were recorded at 25°C using a Watergate W5 pulse sequence for solvent suppression (29). Proton resonances were assigned from the analysis of two-dimensional total correlation spectroscopy and nuclear Overhauser effect spectroscopy experiments, as described previously (20).
RESULTS GAG/CPP binding affinity can be controlled by the number of Trp residues in peptide sequences We first examined the capacity of penetratin, (W48F)penetratin mutant, R9, TAT, R6W3, R6L3, and R10W6, peptides (Table 1) to bind carbohydrates in vitro. We analyzed the interaction of CPPs with HI and 4 different classes of CSs (the most prevalent GAG on the cell surface): monosulfated CSA and CSC and disulfated CSD and CSE (Fig. 1). The interaction of these peptides with the different polysaccharides in solution was analyzed by ITC experiments (Table 2). The ionic strength experimental conditions ensure that the negative charges along the polymer chains were fully screened by the positive ion salts. The presence of EDTA, a divalent cation chelating agent, did not significantly affect the binding parameters between peptides and GAGs. At the saturation conditions, the dissociation constants were in the nanomolar to submicromolar range for all peptides. The affinity ranges for all peptides were mostly in order according to the sulfate content of those carbohydrates (Fig. 1); that is, HI, CSE 3
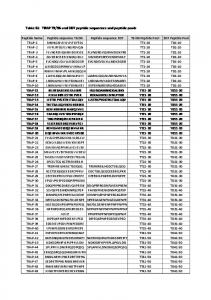
TABLE 2. Thermodynamic parameters of GAG binding to CPPs determined by ITC at 37°C
Peptide and Z
GAG
Kd (nM)
⌬H° (kcal/mol GAG)
T⌬S° (kJ/mol GAG)
Stoichiometry (peptide/GAG)
R9, ⫹9
HI CSE CSD CSC CSA HI CSE CSD CSC CSA HI CSE CSD CSC CSA HI CSE CSD CSC CSA HI CSE CSD CSC CSA HI CSE CSD CSC CSA HI CSE CSD CSC CSA
8.7 ⫾ 2.6 1.0 ⫾ 0.4 180 ⫾ 28 230 ⫾ 9 39 ⫾ 15 36 ⫾ 22 8.8 ⫾ 8.5 53 ⫾ 36 26 ⫾ 4 91 93 ⫾ 20 36 ⫾ 5 115 ⫾ 69 59 ⫾ 44 280 7.2 ⫾ 5.4 16 ⫾ 6 51 ⫾ 27 35 ⫾ 13 38 ⫾ 6 22 ND 170 290 ND 1.5 ND 12 8 ND 76 ⫾ 6 62 210 ⫾ 115 NI NI
⫺60 ⫾ 6 ⫺123 ⫾ 15 ⫺48 ⫺96 ⫾ 44 ⫺42 ⫾ 22 ⫺58 ⫾ 6 ⫺120 ⫾ 31 ⫺39 ⫺56 ⫾ 7 ⫺40 ⫺83 ⫾ 9 ⫺268 ⫾ 53 ⫺96 ⫾ 6 ⫺200 ⫾ 148 ⫺107 ⫺116 ⫾ 2 ⫺427 ⫾ 72 ⫺163 ⫾ 20 ⫺253 ⫾ 37 ⫺171 ⫾ 5 ⫺83 ND ⫺110 ⫺154 ND ⫺124 ND ⫺172 ⫺340 ND ⫺64 ⫾ 3 ⫺145 ⫺46 ⫾ 10 NI NI
⫺48 ⫾ 6 ⫺110 ⫾ 15 ⫺38 ⫺87 ⫾ 44 ⫺32 ⫾ 22 ⫺46 ⫾ 6 ⫺108 ⫾ 32 ⫺28 ⫺45 ⫾ 7 ⫺30 ⫺73 ⫾ 9 ⫺258 ⫾ 53 ⫺87 ⫾ 7 ⫺190 ⫾ 148 ⫺98 ⫺104 ⫾ 1 ⫺416 ⫾ 72 ⫺152 ⫾ 20 ⫺243 ⫾ 37 ⫺160 ⫾ 5 ⫺73 ND ⫺100 ⫺144 ND ⫺111 ND ⫺160 ⫺328 ND ⫺54 ⫾ 3 ⫺135 ⫺36 ⫾ 10 NI NI
8⫾1 37 ⫾ 4 21 ⫾ 5 48 ⫾ 8 26 ⫾ 1 10 ⫾ 1 43 ⫾ 6 22 ⫾ 1 73 ⫾ 7 31 11 ⫾ 1 41 ⫾ 3 27 ⫾ 8 58 ⫾ 9 43 12 ⫾ 1 75 ⫾ 16 33 ⫾ 2 65 ⫾ 18 35 ⫾ 2 7 ND 25 84 ND 10 ND 14 28 ND 13 58 60 ⫾ 1 NI NI
TAT, ⫹8
Penetratin, ⫹7
R6W3, ⫹6
(W48F)-penetratin, ⫹7
R10W6, ⫹10
R6L3, ⫹6
ND, not determined; NI, no interaction in the studied conditions.
⬎ CSD, CSC, CSA (Table 2), with the exception of R6L3. Under the same concentration conditions as for the other CPPs, 50 –100 M R6L3 and 10 –15 M CSA or CSC, no heat was released. To identify the molecular rationale behind those data, we plotted the Kd data according to the number of positive charges (Fig. 2A), the number of Arg or Lys residues, the number of aromatic (Trp, Phe, or Tyr) residues, or the number of Trp residues. A correlation was found only for the plot of Kd according to the number of Trp residues. We observed a linear relationship between the free energy values (⌬G) and the number of Trp residues nW, meaning that the affinity is increasing linearly with the number of Trp residues (Fig. 2B), for HI as Kd ⫽ 55e⫺0.6(nW) and for CSD as Kd ⫽ 250e⫺0.5(nW). Mechanism of binding to GAG relies on Trp residues in CPP sequences The interaction of CPPs with carbohydrates has been studied so far with just a few peptide sequences and a 4
Vol. 27
February 2013
few types of GAGs. The interactions of TAT, oligoarginine, and penetratin with HI, of TAT and oligoarginine with HS, and of TAT with CSB have been reported independently (16, 17, 30, 31). Favorable binding enthalpy The interaction was found to be exothermic for all GAG-peptide complexes (Table 2). Surprisingly, with the exception of TAT and oligoarginine, the interaction of CPPs with HI gave the weakest binding enthalpy, whereas the order of binding enthalpy for CSs was nearly the same as that for binding affinity; that is, CSE, CSC ⬎ CSD, CSA. Thus, apart from HI, the binding enthalpy was stronger for GAGs with higher molecular weight, showing that not only the charge of the GAGs but also the size is a crucial binding parameter; this was confirmed by the absence of interaction of penetratin with SA monomers (up to 5 mM concentration). Among peptide sequences, those containing Trp residues [penetratin, (W48F)-penetratin, R6W3, and R10W6] had stronger binding enthalpy than other basic pep-
The FASEB Journal 䡠 www.fasebj.org
BECHARA ET AL.
that contain hydrophobic Trp residues, compared with other basic CPPs. Stoichiometry of GAG/CPP complexes attains electric neutrality
Figure 2. A, C, E) Binding affinity (A), binding enthalpy (C) and binding entropy (E) according to the number of positive charges (number of Arg/Lys) in the CPPs for the GAGs investigated. B, D, F) Plots of the binding affinity (B), the binding enthalpy (D), and the binding entropy (F) according to the number of Trp residues in peptide sequences; regression coefficients (R2) were all between 0.85 and 0.99.
tides (R9, TAT, and R6L3), irrespective of the type of GAG. In this manner, it was found that the binding enthalpy did not vary significantly with the number of positive charge (Fig. 2C), but became stronger with an increased number of Trp residues (Fig. 2D): the more Trp residues present in the peptide, the more favorable was the binding enthalpy. The binding enthalpy increased from 2- to 4.5-fold with 1 and 6 Trp residues, respectively, compared with peptides that do not contain Trp residues (R9, Tat, and R6L3). The improvement in binding enthalpy observed for peptides containing Trp, was reflected in better affinity (Fig. 2C), because the enthalpy gain dominated unfavorable entropy (Table 2 and Fig. 2D). Unfavorable binding entropy As for binding enthalpy, the unfavorable binding entropy was not dependent on the number of positive charges, because the R9, TAT, and R6L3 interaction gave similar entropy values (Fig. 2E). However, unfavorable entropy increased linearly with increasing numbers of Trp residues in the peptide sequence (Fig. 2F). Thus, greater unfavorable conformational entropy (more conformational penalties or fewer degrees of freedom) arose from the binding to GAGs of peptides CATIONIC CELL-PENETRATING PEPTIDE INTERNALIZATION
The differences in binding enthalpy and entropy were more significant with CSC and CSE than with the other polysaccharides. This observation demonstrated that the degree of sulfation and size of the polysaccharide were crucial parameters, which determined the binding energy of CPPs. On the basis of the molecular weight and sulfate content of the CS and HI polysaccharides, the total negative charge per molecule was calculated (Fig. 1). Taking into account the net positive charge of each peptide and the stoichiometry of their binding to GAGs (Table 2), the interaction led to CPP-GAG neutralized charge complexes with all polysaccharides (Supplemental Table S1). However, the interaction of CPPs with GAGs not only occurred through ion pairs but also the peptide sequence, the length, and the position or degree of sulfation in the polysaccharide were determinant binding factors. In fact, the binding affinity increased as a function of sulfate content and chain length of GAGs (Fig. 2A). Notably, no interaction of penetratin with SA alone (up to 5 mM) could be detected. Trp-rich CPPs adopt predominantly a -strand conformation in complex with GAGs Although usually unstructured in solution, CPPs tend to adopt a well-defined secondary structure when binding to lipid membranes (32). As for the interaction with polysaccharides, it has been shown that TAT peptide does not undergo conformational changes in the presence of HS (30). On the other hand, different studies demonstrated that HI binding peptides adopt an ␣-helical structure when bound to HI (and other polyelectrolytes; refs. 33, 34). Here, we used CD spectroscopy and NMR to determine the secondary structure of cationic peptides in the presence of carbohydrates. Based on the stoichiometry of binding calculated in the ITC experiments, we tested different peptide/HI ratios to have the peptide either saturated or not saturated with GAGs (Table 3). Except for R6W3 and R10W6, all the peptides had a random coil structure in buffer solution. R6W3, on the other hand, had partial helical content that was previously characterized, with a minimum shifted to 202 nm and another shouldering at 220 nm. This peculiar signature was ascribed to the presence of Trp residues (20). On the other hand, R10W6 could accommodate ␣-helical and -strand conformations in solution. Conformational differences among peptides were unveiled when bound in complex with HI (Table 3). R9, TAT, and R6L3 were partially structured into an ␣ helix (Supplemental Fig. S1). On the other hand, R6W3, R10W6, penetratin, and (W48F)-penetratin could adopt a -strand structure principally (minimum at 213 nm 5
TABLE 3. Secondary structure content of the CPPs in phosphate buffer (pH 7.4) or in the presence of HI at saturating peptide conditions
CPP
Penetratin Buffer HI (W48F)-penetratin Buffer HI R10W6 Buffer HI R6W3 Buffer HI R6L3 Buffer HI R9 Buffer HI TAT Buffer HI
␣-Helix (%)
Random coil (%)
Type II helix (%)
-Sheet (%)
1 3
57 69
42 0
0 28
0 31
58 51
42 5
0 13
21 8
65 55
0 0
14 37
18 25
71 48
11 0
0 27
0 47
77 35
23 0
0 18
0 18
83 70
17 12
0 0
0 13
92 78
8 0
0 9
Experimental error is estimated to be ⫾5% (28).
and maximum at 193 nm; Supplemental Fig. S1). Thus, peptides with Trp residues could adopt predominantly a higher-order -strand conformation in interaction with HI, better than that of the other basic peptides, which remained unstructured or adopted an ␣-helical conformation. It should be noted that on addition of the peptides to HI, a fine precipitate formed in the solution. This might be due to the aggregation phe-
nomena that take place with CPP-HI binding, through formation of -sheet aggregates (13, 16, 17). Light scattering due to the particulate nature of the solution might have interfered with the CD experiment, but, nonetheless, the increase in ordered structure content came at the expense of other secondary structural features. The interaction of CPPs with GAGs was further examined by 1H NMR spectroscopy. Taken as model peptides, R6L3 and R6W3 were titrated with increasing concentrations of HI-derived oligosaccharides. The titrations with HI tetrasaccharide DP4 (Fig. 3) show very different behaviors for R6L3 and R6W3 peptides. In the case of R6L3 (Fig. 3A), the amide resonances undergo gradual chemical shift perturbations on titration, indicating an interaction occurring on a fast exchange regimen on the NMR time scale. No significant broadening is observed, suggesting that the interaction does not involve the formation of high-molecular-weight species. The amide protons of residues 3–9 exhibit an average chemical shift variation of 0.17 ppm that can be ascribed to direct interaction with heparin tetrasaccharide or peptide folding. Notably, the H␣ protons also show an average upfield shift of 0.07 ppm, which supports a helical folding of the R6L3 peptide on complex formation. In contrast, the proton resonances of R6W3 peptide show a gradual decrease in intensity without any changes in chemical shifts or linewidths as the DP4 concentration is increased, whereas no peptide resonances can be observed beyond a 1:1 peptide/DP4 ratio (Fig. 3B). The addition of 1 M NaCl in the sample leads to peptide resonances reappearing, with chemical shifts similar to that of the peptide in the absence of DP4 ligand (Fig. 3C). This observation demonstrates that an NMR-invisible peptide-GAG complex is formed during the titration, which can be dissociated at high
Figure 3. NMR titration of R6L3 and R6W3 peptides with HI tetrasaccharide DP4. A) Amide region of the 1-dimensional 1H NMR spectra of R6L3 as a function of added DP4 (peptide/DP4 ratios of 1:0 to 1:5) B, C) Amide region of the 1-dimensional 1H NMR spectra of R6W3 as a function of added DP4 (peptide/DP4 ratios of 1:0 to 1:1) in the presence of 150 mM NaCl (B) or 1 M NaCl (C).
6
Vol. 27
February 2013
The FASEB Journal 䡠 www.fasebj.org
BECHARA ET AL.
ionic strength. From the titration, it can be calculated that the interaction involves a binding of 0.7 peptide per disaccharide unit and leads to electrostatic neutrality of the complex. Therefore, as opposed to R6L3, the interaction of R6W3 with HI saccharides leads to the high-molecular-weight aggregates consistent with -sheet formation, which are too large to be observed by solution-state NMR. Trp-rich peptides form stable and large aggregates in complex with GAGs Because precipitates were observed in saturated peptide-GAG complexes, we further explored this possibility by turbidity and differential light-scattering measures and phase-contrast microscopy. It was previously reported that conformational transition of penetratin from an ␣ helix to a  strand could be associated with aggregation on lipid binding (35, 36). Turbidity measurements of saturated GAG-CPP ITC samples showed that there were no significant or rational differences in absorbance among peptides and GAGs (not shown). Obviously, all peptides induced supramolecular assemblies of GAGs in solution, with the exception of R6L3 with CSA and CSC, as expected from the absence of interaction found between the peptide and those carbohydrate molecules in ITC experiments. Thus, in principle, all peptide-GAG complexes could form aggregated molecular species. Dynamic light scattering (DLS) measurements did not help to further decipher the mechanism because the supramolecular assemblies between GAGs and peptides were too polydispersed. In addition, accurate DLS measurement requires particles to be randomly diffusing. This places the upper size limit of DLS analysis as the point at which sedimentation of the particle dominates diffusion, and the upper size limit was obviously reached in the case of the CPP-GAG complexes. Phasecontrast microscopy imaging was thus used to observe the number and size of aggregates formed between HI, monosulfated CSD, disulfated CSC, and CPPs (Supplemental Fig. S2). Trp-rich peptides systematically induced the formation of numerous, large, and stable GAG aggregates (as can be seen in Supplemental Fig. S2). The situation was quite different for the other basic peptides (R9, TAT, and R6L3), for which few or no aggregates were observed, with the exception of HI. In this latter case, TAT and R6L3 caused the formation of many small aggregates, whereas for other peptides, including R9, larger aggregates were stably formed. This observation highlights the fact that HI behaves differently from the other GAGs and is not representative of the interactions of all types of GAGs with CPPs. The higher density of negative charges in HI might be the reason for such differences in the aggregation behavior of this carbohydrate compared with that of others. Thus, these results demonstrated that stable aggregates have been formed between the Trp-rich peptides and the different types of GAGs. On the other hand, for CATIONIC CELL-PENETRATING PEPTIDE INTERNALIZATION
peptides that do not contain Trp residues, either the aggregates were formed but transiently so that they could not be observed, or they were simply not induced during peptide-GAG interaction. Trp-rich peptides internalize efficiently through cell surface GAG clustering We further explored the role of cell surface carbohydrates in the interaction of Trp-rich peptides with plasma membrane. Plasma membrane glycoconjugates form a pool of negative charges on the cell surface that bind the positively charged CPPs, thus facilitating their interaction with the cell (10). We used 3 CHO cell lines that express all types of GAGs (WT), no CS and HS (GAGneg), or no SAs (SAneg). Membrane translocation and endocytosis depending on GAG clustering are observed for CPPs The internalization of cell-penetrating peptides occurs at both 37 and 4°C. It is generally assumed that all translocation and endocytic processes are active at 37°C, whereas endocytic pathways are inhibited at 4°C. Therefore, the amount of internalized peptide via direct membrane translocation pathways can be estimated from the comparison of internalization at 37 and 4°C. This amount might be underestimated because the gel-phase state of the lipid membrane at 4°C might hinder the translocation process. Nevertheless, we chose to modify temperature to distinguish between translocation and endocytic pathways rather than use endocytosis inhibitors because these inhibitors have negative side effects in internalization studies (37). At low micromolar extracellular peptide concentrations and at 37°C, the amount of internalized peptide was similar in all three cell types for both penetratin and R6W3 peptides. This result was also observed previously with a penetratin analog and was related to membrane translocation pathways (12). When the peptide concentration is increased, a significantly higher uptake in SAneg and WT cells than in GAGneg cells was measured. This finding supports previous results showing that a minimum amount of bound CPP is necessary to trigger cooperative GAG clustering and internalization (12, 13, 16, 17). Notably, the internalization was significantly more efficient in SAneg cells than in WT cells (⬎5 M extracellular peptide concentration). Thus, at the cell surface, SAs are inert CPP binders that trap CPPs and prevent them from interacting with other membrane partners, such as GAGs and plasma membrane lipids involved in internalization processes, as described previously (7). Internalization was also visualized by confocal microscopy, at 37°C in the three cell lines. Because it was not possible to observe any differences in peptide quantity among cell lines with this technique, experiments are shown for penetratin only (Supplemental Fig. S4). At 4°C, internalization occurred with similar amounts irrespective of the cell surface carbohydrate composition 7
Figure 4. A, B) Quantification of the internalized peptides R6W3 (A) and penetratin (B) in 106 WT, GAGneg, and SAneg cells at 37 and 4°C. Error bars were obtained from n ⬎ 4 independent experiments. C) Plots of the amount of internalized peptides (logarithmic scale) in WT and GAGneg cells at 37 and 4°C, according to the number of positively charged amino acids [n(⫹)], the number of total aromatic residues [n(W,Y,F)], the number of Trp residues [n(W)], and the number of Arg residues [n(Arg)].
and the extracellular peptide concentration (Fig. 4A, B for R6W3 and penetratin intracellular amounts, respectively, and Supplemental Fig. S4 for intracellular imaging of penetratin). As for 37°C, one should note that R6W3 was internalized more efficiently than penetratin. Finally, these data indicate that these peptides enter via direct translocation at low extracellular concentrations and that both GAG-dependent endocytosis and direct translocation occur at higher concentrations, as described previously for a penetratin analog (12). Efficiency of internalization can be controlled by Trp residues within CPP sequences To further evaluate the effect of CPP sequences on internalization efficiency, the amount of peptide internalized at a saturating (10 M) extracellular peptide concentration was plotted for each peptide, taking into account the number of positive charges and the number of aromatic (F, Y, W) or Arg (R) or Trp (W) residues (Fig. 4C). The internalization data were obtained with WT and GAGneg cells at 37 and 4°C. The highest efficiency of internalization was obtained for R9 and R6W3, which enter cells in similar amounts. This result shows that basic residues in CPPs can be partially replaced by tryptophans, maintaining the same efficiency of uptake as was also observed with pseudopeptide carriers (21). For the other peptide sequences, it is clear that in the two cell lines, regardless of the temperature, the amounts of internalized peptides correlated with the number of Trp residues (and not the total number of aromatic residues; Fig. 4C). The internalization curve fits showed decreased efficiency of internalization in the absence of GAG or at 4°C compared with that f wild-type cells at 37°C. However, the curve fits were shifted parallel lines (Fig. 4C), an observation demonstrating that crucial steps in the mechanism of internalization are not influenced by temperature. 8
Vol. 27
February 2013
DISCUSSION Our data show that introducing Trp residues into the CPP sequence can enhance the entry efficiency of CPPs as recently reported (38). The role of Trp in efficient internalization of peptides is observed not only for direct translocation, as already described for penetratin (39 – 41), but also for endocytosis through GAGs, although these peptides are not cytotoxic (42– 44). Positive charges in the sequence of CPPs are necessary to interact with negatively charged GAGs and phospholipids, but the additional presence of one or more Trp residues governs the binding properties of the peptides. In addition, our results show that Trp-rich peptides can adopt a -strand structure in contact with GAGs, which lead to higher-order complexes and probably form -sheet stable and large aggregates. These results were consistent with the thermodynamic data. A strong binding enthalpy contributed in a major way to the favorable free energy change that was measured for all CPPs. Thus, strong ion pairs, hydrogen bonds, and van der Waals contacts dominate the unfavorable enthalpy associated with the desolvation of polar groups in GAGs and peptides. However, Trp-rich peptides had stronger binding enthalpy and higher entropy loss compared with other basic sequences, which can be explained by the fact that hydrogen bonds of -strands are considered slightly stronger than those found in ␣-helices. With regard to entropy of binding, two major factors contribute to it: desolvation and conformational entropy changes. The desolvation entropy should be favorable because it originates from the release of water molecules on interaction between the peptide and GAGs. Favorable desolvation entropy should arise from the binding energy of hydrophobic groups. However, the conformational entropy change should be unfavorable, because the binding process involves the loss of conformational degrees of freedom for both the peptide and GAGs. Thus, the stronger hydrogen bonds in the Trp-containing peptide-GAG interaction lead to
The FASEB Journal 䡠 www.fasebj.org
BECHARA ET AL.
strong unfavorable conformational entropy that should dominate the favorable desolvation entropy. Notably, Ye et al. (45) reported in live cells that the secondary structure of penetratin (used at high concentration, 20 M) was found to be mainly random coil and  strand in the cytoplasm and  sheet in the nucleus. With regard to the molecular mechanisms of CPPGAG interactions, the first binding step should involve ion-pair formation between all basic peptides and GAGs. Arg and Trp residues could contact the sugar units either by electrostatic and bidentate hydrogen bond interactions with the sulfates or by hydrophobic interactions to the sugar rings, respectively. Trp residues may also bind to sulfate groups of GAGs (46) through -anion interactions (47– 49). In addition, it is well-known that Trp and Arg side chains from two different peptide  strands can also interact through -cation noncovalent bonds and evoke self-assembly of peptides, which should lead to the formation of -sheet aggregates in complex with GAGs. Finally, -cation interactions between Trp and Arg residues can tune the pKa of the guanidinium side chain, increasing the abundance of the protonated form of arginyl residues. Together these interactions increase both the binding enthalpy and the unfavorable binding entropy of peptide-GAG complexes. In the absence of Trp, Arg, more so than Lys, mediated the binding properties of CPPs, which better fit ␣-helical conformational structures. This was not surprising because Arg is a better HI binder than Lys (46, 50) through the formation of higher-energy bidentate hydrogen bondings. These peptide helices do not form stable and large aggregates as do Trp-rich peptides. However, with a minimum number of 9 Arg residues, a level of internalization similar to that of a peptide containing 6 Arg and 3 Trp residues was recovered, suggesting that ␣-helical peptides with a higher density of positive charges could also transiently but efficiently aggregate GAGs and internalize (17). Thus, Trp-rich basic peptides are more prone than other CPPs to form stable clusters of GAGs, when GAG clustering is a key step for subsequent endocytosis (10, 51, 52). In the case studied herein, this property is not related to the primary or secondary amphipathic structure of the peptides, because R6L3 and R6W3, with similar amphipathic profiles, behaved differently in terms of interaction with GAGs and internalization. However, the position of Trp within the peptide sequence can be an important issue (38). In the absence of Trp residues, the number of Arg residues more so than the number of positive charges, regulates the efficient internalization of CPPs into cells. In this manner, a minimum number of 6 Arg residues seems requisite for entry of conventional peptide sequences into cells, as reported previously (53, 54). Thus, our data highlight the importance of Trp and Arg residues in the binding, structuring, and internalization of some CPP sequences through GAG-dependent endocytosis but also through translocation processes. However, this CATIONIC CELL-PENETRATING PEPTIDE INTERNALIZATION
result does not exclude the possibility that other amino acids may be important for GAG interaction (55). Finally, the fact that the amounts of internalized peptides are linearly proportional to the number of Trp residues in a CPP sequence, regardless of the cell surface (the absence or the presence of GAGs) and temperature (37 or 4°C) strongly suggests a shared first step in the mechanism for both internalization pathways: interaction with GAGs for endocytosis and with phospholipids for membrane perturbation/translocation. It was previously reported that penetratin can adopt a -strand conformation in complex with acidic (negatively charged) phospholipids and that the charge density is a crucial parameter in the translocation process (35, 36, 56, 57). It is also shown herein that the negative charge alone is not sufficient to allow for CPP binding, because no interaction of penetratin with SA alone could be detected. These results are consistent with those from Ziegler and Seelig (16), who showed that the cationic charge alone is not sufficient to provide binding to GAGs. Indeed, these researchers reported a tight binding of nona-arginine but not of the isolated arginine amino acid for HI, although studied at identical arginine monomer concentrations (1.2 mM for nona-arginine and 10 mM for Arg; ref. 16). This result demonstrates that interactions at low nanomolar concentrations require a minimum charge density to fulfill the polyelectrolyte theory, as already pointed out (58). Thus, we propose, as a first step for internalization of cationic CPP, clustering of negatively charged molecules, either lipids or carbohydrates, at the cell surface, triggered differently by peptides containing Trp, which further leads to translocation or endocytosis, respectively (Scheme 1). For the mechanism of endocytosis, an energy- and temperature-dependent process, invagination or ruffling of the plasma membrane, could occur, a process assisted by GTPase activity and actin dynamics (59). For membrane translocation, different types of membrane perturbation have been described so far: inverted micelles (60), membrane tubulation (61), transient pore-formation/ membrane repair (62), or permeant interface between segregated gel-like and more fluid membrane domains (63). A previous study has shown that the process of translocation or membrane crossing of a penetratin analog, was limited in time and reached a plateau in kinetics experiments (12), a result that favors the hypothesis of a coentry of CPP with negatively charged glycolipids or lipids. In summary, we found that the entry of peptides into cells can be rationally controlled by the presence of Arg and Trp in the CPP sequence. The ␣ helix and  strand, previously described as being essential secondary structures for interaction with HI (64), do not have the same efficiency in inducing GAG clustering;  strands are better inducers of stable GAG aggregates, which can be related to their better internalization efficiency into cells than ␣ helices. This finding is beyond the scope of CPPs and encom9
Scheme 1. Mechanisms of internalization (translocation and endocytosis) of CPPs. Unstructured in solution, positively charged CPPs interact with negatively charged polymers (GAGs and phospholipids) at the cell surface. The interaction between CPPs and negatively charged polymers leads to a -strand conformation of Trp-containing peptides and to an ␣-helical structure for other basic peptide sequences. Interactions with CPPs trigger the clustering of the negatively charged molecules, which form stable aggregated -sheet complexes with Trp-containing peptides and might lead to transient aggregates for basic ␣-helical peptides. Then, membrane ruffling or invagination of the plasma membrane evokes endocytosis of CPP/GAG complexes, whereas membrane destabilization/perturbation of the lipid membrane leads to direct translocation of the peptides into cells.
passes the role of Trp residues in the interaction of any signaling peptide or protein, with either secreted carbohydrates or those attached to the cell surface. Notably, in prokaryotes, antimicrobial activity could be enhanced with Trp end-tagged antimicrobial peptide sequences (65). In this latter case, the Trptagged antimicrobial peptide sequences had increased potency not only to bind the negatively charged lipopolysaccharide but also to evoke wall rupture in bacteria or a model lipid vesicle. As for eukaryotes, it has been shown that the protein PDX-1, a protein containing an antennapedia-like protein transduction domain, has two Trp residues important for the internalization of the protein (66, 67). Conserved Trp residues are also found in homeodomains of homeobox proteins (68), which can traffic from cell to cell (69, 70). The acidic fibroblast growth factor protein also contains a Trp residue in a -strand domain that interacts with HI (71). Thus, as a general rule, interaction and signaling properties of endogenous basic peptides and proteins have to be reexamined in the light of the potential strong interactions between Trp residues and carbohydrates at the cell surface. Support for this research was provided the Université Pierre et Marie Curie (UPMC; Université Paris 6), by ANRBLAN2010-ParaHP (postdoctoral position for M.P.), the Centre National de la Recherche Scientifique (CNRS), and the 10
Vol. 27
February 2013
French Ministère de l’Enseignement Supérieur et de la Recherche (Ph.D. fellowship for C.B.). Y.Z. was supported by the U.S. National Science Foundation (grant CHE-0755225, for a fellowship sponsored by the University of Florida).
REFERENCES 1. 2.
3. 4. 5. 6.
7.
8. 9.
Futaki, S., Nakase, I., Tadokoro, A., Takeuchi, T., and Jones, A. T. (2007) Arginine-rich peptides and their internalization mechanisms. Biochem. Soc. Trans. 35, 784 –787 Alves, I. D., Jiao, C. Y., Aubry, S., Aussedat, B., Burlina, F., Chassaing, G., and Sagan, S. (2010) Cell biology meets biophysics to unveil the different mechanisms of penetratin internalization in cells. Biochim. Biophys. Acta 1798, 2231–2239 Lindgren, M., and Langel, U. (2011) Classes and prediction of cell-penetrating peptides. Metab. Mol. Biol. 683, 3–19 Jones, L. S., Yazzie, B., and Middaugh, C. R. (2004) Polyanions and the proteome. Mol. Cell. Proteomics 3, 746 –769 Varki, A. (2007) Glycan-based interactions involving vertebrate sialic-acid-recognizing proteins. Nature 446, 1023–1029 Joliot, A. H., Triller, A., Volovitch, M., Pernelle, C., and Prochiantz, A. (1991) ␣-2,8-Polysialic acid is the neuronal surface receptor of antennapedia homeobox peptide. New Biol. 3, 1121–1134 Alves, I. D., Bechara, C., Walrant, A., Zaltsman, Y., Jiao, C. Y., and Sagan, S. (2011) Relationships between membrane binding, affinity and cell internalization efficacy of a cell-penetrating peptide: penetratin as a case study. PloS One 6, e24096 Zimmermann, P., and David, G. (1999) The syndecans, tuners of transmembrane signaling. FASEB J. 13(Suppl), S91⫺S100 Sasisekharan, R., and Venkataraman, G. (2000) Heparin and heparan sulfate: biosynthesis, structure and function. Curr. Opin. Chem. Biol. 4, 626 –631
The FASEB Journal 䡠 www.fasebj.org
BECHARA ET AL.
10.
11. 12.
13.
14.
15.
16. 17. 18.
19.
20.
21.
22.
23. 24.
25.
26. 27.
28.
Belting, M., Mani, K., Jonsson, M., Cheng, F., Sandgren, S., Jonsson, S., Ding, K., Delcros, J. G., and Fransson, L. A. (2003) Glypican-1 is a vehicle for polyamine uptake in mammalian cells: a pivotal role for nitrosothiol-derived nitric oxide. J. Biol. Chem. 278, 47181–47189 Belting, M. (2003) Heparan sulfate proteoglycan as a plasma membrane carrier. Trends Biochem. Sci. 28, 145–151 Jiao, C. Y., Delaroche, D., Burlina, F., Alves, I. D., Chassaing, G., and Sagan, S. (2009) Translocation and endocytosis for cellpenetrating peptide internalization. J. Biol. Chem. 284, 33957– 33965 Ziegler, A., and Seelig, J. (2011) Contributions of glycosaminoglycan binding and clustering to the biological uptake of the nonamphipathic cell-penetrating peptide WR9. Biochemistry 50, 4650 –4664 Ram, N., Aroui, S., Jaumain, E., Bichraoui, H., Mabrouk, K., Ronjat, M., Lortat-Jacob, H., and De Waard, M. (2008) Direct peptide interaction with surface glycosaminoglycans contributes to the cell penetration of maurocalcine. J. Biol. Chem. 283, 24274 –24284 Letoha, T., Keller-Pinter, A., Kusz, E., Kolozsi, C., Bozso, Z., Toth, G., Vizler, C., Olah, Z., and Szilak, L. (2010) Cellpenetrating peptide exploited syndecans. Biochim. Biophys. Acta 1798, 2258 –2265 Ziegler, A., and Seelig, J. (2008) Binding and clustering of glycosaminoglycans: a common property of mono- and multivalent cell-penetrating compounds. Biophys. J. 94, 2142–2149 Rullo, A., Qian, J., and Nitz, M. (2011) Peptide-glycosaminoglycan cluster formation involving cell penetrating peptides. Biopolymers 95, 722–731 Yang, H., Liu, S., Cai, H., Wan, L., Li, S., Li, Y., Cheng, J., and Lu, X. (2010) Chondroitin sulfate as a molecular portal that preferentially mediates the apoptotic killing of tumor cells by penetratin-directed mitochondria-disrupting peptides. J. Biol. Chem. 285, 25666 –25676 Wittrup, A., Zhang, S. H., ten Dam, G. B., van Kuppevelt, T. H., Bengtson, P., Johansson, M., Welch, J., Morgelin, M., and Belting, M. (2009) ScFv antibody-induced translocation of cellsurface heparan sulfate proteoglycan to endocytic vesicles: evidence for heparan sulfate epitope specificity and role of both syndecan and glypican. J. Biol. Chem. 284, 32959 –32967 Walrant, A., Correia, I., Jiao, C. Y., Lequin, O., Bent, E. H., Goasdoue, N., Lacombe, C., Chassaing, G., Sagan, S., and Alves, I. D. (2011) Different membrane behaviour and cellular uptake of three basic arginine-rich peptides. Biochim. Biophys. Acta 1808, 382–393 Aussedat, B., Sagan, S., Chassaing, G., Bolbach, G., and Burlina, F. (2006) Quantification of the efficiency of cargo delivery by peptidic and pseudo-peptidic Trojan carriers using MALDI-TOF mass spectrometry. Biochim. Biophys. Acta 1758, 375–383 Delaroche, D., Aussedat, B., Aubry, S., Chassaing, G., Burlina, F., Clodic, G., Bolbach, G., Lavielle, S., and Sagan, S. (2007) Tracking a new cell-penetrating (W/R) nonapeptide, through an enzyme-stable mass spectrometry reporter tag. Anal. Chem. 79, 1932–1938 Derossi, D., Chassaing, G., and Prochiantz, A. (1998) Trojan peptides: the penetratin system for intracellular delivery. Trends Cell Biol. 8, 84 –87 Burlina, F., Sagan, S., Bolbach, G., and Chassaing, G. (2005) Quantification of the cellular uptake of cell-penetrating peptides by MALDI-TOF mass spectrometry. Angew. Chem. Int. Ed. 44, 4244 –4247 Burlina, F., Sagan, S., Bolbach, G., and Chassaing, G. (2006) A direct approach to quantification of the cellular uptake of cell-penetrating peptides using MALDI-TOF mass spectrometry. Nat. Protoc. 1, 200 –205 Binder, H., and Lindblom, G. (2003) Charge-dependent translocation of the Trojan peptide penetratin across lipid membranes. Biophys. J. 85, 982–995 Epand, R. M., Epand, R. F., Arnusch, C. J., PapahadjopoulosSternberg, B., Wang, G., and Shai, Y. (2010) Lipid clustering by three homologous arginine-rich antimicrobial peptides is insensitive to amino acid arrangement and induced secondary structure. Biochim. Biophys. Acta 1798, 1272–1280 Jean-Francois, F., Khemtemourian, L., Odaert, B., Castano, S., Grelard, A., Manigand, C., Bathany, K., Metz-Boutigue, M. H., and Dufourc, E. J. (2007) Variability in secondary structure of
CATIONIC CELL-PENETRATING PEPTIDE INTERNALIZATION
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
the antimicrobial peptide Cateslytin in powder, solution, DPC micelles and at the air-water interface. Eur. Biophys. J. 36, 1019 –1027 Liu, M., Mao, X. A., Ye, C., Huang, H., Nicholson, J. K., and Lindon, J. C. (1998) Improved WATERGATE pulse sequences for solvent suppression in NMR spectroscopy. J. Magn. Reson. 132, 125–129 Ziegler, A., and Seelig, J. (2004) Interaction of the protein transduction domain of HIV-1 TAT with heparan sulfate: binding mechanism and thermodynamic parameters. Biophys. J. 86, 254 –263 Goncalves, E., Kitas, E., and Seelig, J. (2005) Binding of oligoarginine to membrane lipids and heparan sulfate: structural and thermodynamic characterization of a cell-penetrating peptide. Biochemistry 44, 2692–2702 Eiriksdottir, E., Konate, K., Langel, U., Divita, G., and Deshayes, S. (2010) Secondary structure of cell-penetrating peptides controls membrane interaction and insertion. Biochim. Biophys. Acta 1798, 1119 –1128 Mulloy, B., Crane, D. T., Drake, A. F., and Davies, D. B. (1996) The interaction between heparin and polylysine: a circular dichroism and molecular modelling study. Braz. J. Med. Biol. Res. 29, 721–729 Jayaraman, G., Wu, C. W., Liu, Y. J., Chien, K. Y., Fang, J. C., and Lyu, P. C. (2000) Binding of a de novo designed peptide to specific glycosaminoglycans. FEBS Lett. 482, 154 –158 Magzoub, M., Eriksson, L. E., and Graslund, A. (2003) Comparison of the interaction, positioning, structure induction and membrane perturbation of cell-penetrating peptides and nontranslocating variants with phospholipid vesicles. Biophys. Chem. 103, 271–288 Persson, D., Thoren, P. E., and Norden, B. (2001) Penetratininduced aggregation and subsequent dissociation of negatively charged phospholipid vesicles. FEBS Lett. 505, 307–312 Vercauteren, D., Vandenbroucke, R. E., Jones, A. T., Rejman, J., Demeester, J., De Smedt, S. C., Sanders, N. N., and Braeckmans, K. (2010) The use of inhibitors to study endocytic pathways of gene carriers: optimization and pitfalls. Mol. Ther. 18, 561–569 Rydberg, H. A., Matson, M., Amand, H. L., Esbjorner, E. K., and Norden, B. (2012) Effects of tryptophan content and backbone spacing on the uptake efficiency of cell-penetrating peptides. Biochemistry 51, 5531–5539 Christiaens, B., Symoens, S., Verheyden, S., Engelborghs, Y., Joliot, A., Prochiantz, A., Vandekerckhove, J., Rosseneu, M., and Vanloo, B. (2002) Tryptophan fluorescence study of the interaction of penetratin peptides with model membranes. Eur. J. Biochem. 269, 2918 –2926 Lindberg, M., Biverstahl, H., Graslund, A., and Maler, L. (2003) Structure and positioning comparison of two variants of penetratin in two different membrane mimicking systems by NMR. Eur. J. Biochem. 270, 3055–3063 Christiaens, B., Grooten, J., Reusens, M., Joliot, A., Goethals, M., Vandekerckhove, J., Prochiantz, A., and Rosseneu, M. (2004) Membrane interaction and cellular internalization of penetratin peptides. Eur. J. Biochem. 271, 1187–1197 Aubry, S., Burlina, F., Dupont, E., Delaroche, D., Joliot, A., Lavielle, S., Chassaing, G., and Sagan, S. (2009) Cell-surface thiols affect cell entry of disulfide-conjugated peptides. FASEB J. 23, 2956 –2967 Delaroche, D., Cantrelle, F. X., Subra, F., Van Heijenoort, C., Guittet, E., Jiao, C. Y., Blanchoin, L., Chassaing, G., Lavielle, S., Auclair, C., and Sagan, S. (2010) Cell-penetrating peptides with intracellular actin-remodeling activity in malignant fibroblasts. J. Biol. Chem. 285, 7712–7721 Lamaziere, A., Burlina, F., Wolf, C., Chassaing, G., Trugnan, G., and Ayala-Sanmartin, J. (2007) Non-metabolic membrane tubulation and permeability induced by bioactive peptides. PloS One 2, e201 Ye, J., Fox, S. A., Cudic, M., Rezler, E. M., Lauer, J. L., Fields, G. B., and Terentis, A. C. (2010) Determination of penetratin secondary structure in live cells with Raman microscopy. J. Am. Chem. Soc. 132, 980 –988 Mascotti, D. P., and Lohman, T. M. (1995) Thermodynamics of charged oligopeptide-heparin interactions. Biochemistry 34, 2908 –2915
11
47. 48. 49.
50.
51. 52.
53.
54.
55.
56.
57.
58. 59.
12
Demeshko, S., Dechert, S., and Meyer, F. (2004) Anion-pi interactions in a carousel copper(II)-triazine complex. J. Am. Chem. Soc. 126, 4508 –4509 Maeda, H., Osuka, A., and Furuta, H. (2004) Anion binding properties of N-confused porphyrins at the peripheral nitrogen. J. Incl. Phenom. Macrocycl. Chem. 49, 33–36 Ito, K., Olsen, S. L., Qiu, W., Deeley, R. G., and Cole, S. P. (2001) Mutation of a single conserved tryptophan in multidrug resistance protein 1 (MRP1/ABCC1) results in loss of drug resistance and selective loss of organic anion transport. J. Biol. Chem. 276, 15616 –15624 Fromm, J. R., Hileman, R. E., Caldwell, E. E., Weiler, J. M., and Linhardt, R. J. (1995) Differences in the interaction of heparin with arginine and lysine and the importance of these basic amino acids in the binding of heparin to acidic fibroblast growth factor. Arch. Biochem. Biophys. 323, 279 –287 Poon, G. M., and Gariepy, J. (2007) Cell-surface proteoglycans as molecular portals for cationic peptide and polymer entry into cells. Biochem. Soc. Trans. 35, 788 –793 Sarrazin, S., Wilson, B., Sly, W. S., Tor, Y., and Esko, J. D. (2010) Guanidinylated neomycin mediates heparan sulfate-dependent transport of active enzymes to lysosomes. Mol. Ther. 18, 1268 – 1274 Wender, P. A., Mitchell, D. J., Pattabiraman, K., Pelkey, E. T., Steinman, L., and Rothbard, J. B. (2000) The design, synthesis, and evaluation of molecules that enable or enhance cellular uptake: peptoid molecular transporters. Proc. Natl. Acad. Sci. U. S. A. 97, 13003–13008 Rothbard, J. B., Garlington, S., Lin, Q., Kirschberg, T., Kreider, E., McGrane, P. L., Wender, P. A., and Khavari, P. A. (2000) Conjugation of arginine oligomers to cyclosporin A facilitates topical delivery and inhibition of inflammation. Nat. Med. 6, 1253–1257 Caldwell, E. E., Nadkarni, V. D., Fromm, J. R., Linhardt, R. J., and Weiler, J. M. (1996) Importance of specific amino acids in protein binding sites for heparin and heparan sulfate. Int. J. Biochem. Cell Biol. 28, 203–216 Magzoub, M., Eriksson, L. E., and Graslund, A. (2002) Conformational states of the cell-penetrating peptide penetratin when interacting with phospholipid vesicles: effects of surface charge and peptide concentration. Biochim. Biophys. Acta 1563, 53–63 Bellet-Amalric, E., Blaudez, D., Desbat, B., Graner, F., Gauthier, F., and Renault, A. (2000) Interaction of the third helix of Antennapedia homeodomain and a phospholipid monolayer, studied by ellipsometry and PM-IRRAS at the air-water interface. Biochim. Biophys. Acta 1467, 131–143 Mitchell, D. J., Kim, D. T., Steinman, L., Fathman, C. G., and Rothbard, J. B. (2000) Polyarginine enters cells more efficiently than other polycationic homopolymers. J. Pept. Res. 56, 318 –325 Gerbal-Chaloin, S., Gondeau, C., Aldrian-Herrada, G., Heitz, F., Gauthier-Rouviere, C., and Divita, G. (2007) First step of the
Vol. 27
February 2013
60.
61.
62.
63.
64.
65.
66.
67.
68. 69.
70.
71.
cell-penetrating peptide mechanism involves Rac1 GTPase-dependent actin-network remodelling. Biol. Cell 99, 223–238 Derossi, D., Calvet, S., Trembleau, A., Brunissen, A., Chassaing, G., and Prochiantz, A. (1996) Cell internalization of the third helix of the Antennapedia homeodomain is receptor-independent. J. Biol. Chem. 271, 18188 –18193 Lamaziere, A., Chassaing, G., Trugnan, G., and Ayala-Sanmartin, J. (2009) Tubular structures in heterogeneous membranes induced by the cell penetrating peptide penetratin. Commun. Integr. Biol. 2, 223–224 Palm-Apergi, C., Lorents, A., Padari, K., Pooga, M., and Hallbrink, M. (2009) The membrane repair response masks membrane disturbances caused by cell-penetrating peptide uptake. FASEB J. 23, 214 –223 Verdurmen, W. P., Thanos, M., Ruttekolk, I. R., Gulbins, E., and Brock, R. (2010) Cationic cell-penetrating peptides induce ceramide formation via acid sphingomyelinase: implications for uptake. J. Control. Release 147, 171–179 Margalit, H., Fischer, N., and Ben-Sasson, S. A. (1993) Comparative analysis of structurally defined heparin binding sequences reveals a distinct spatial distribution of basic residues. J. Biol. Chem. 268, 19228 –19231 Pasupuleti, M., Chalupka, A., Morgelin, M., Schmidtchen, A., and Malmsten, M. (2009) Tryptophan end-tagging of antimicrobial peptides for increased potency against Pseudomonas aeruginosa. Biochim. Biophys. Acta 1790, 800 –808 Noguchi, H., Kaneto, H., Weir, G. C., and Bonner-Weir, S. (2003) PDX-1 protein containing its own antennapedia-like protein transduction domain can transduce pancreatic duct and islet cells. Diabetes 52, 1732–1737 Ueda, M., Matsumoto, S., Hayashi, S., Kobayashi, N., and Noguchi, H. (2008) Cell surface heparan sulfate proteoglycans mediate the internalization of PDX-1 protein. Cell Transplant. 17, 91–97 Merabet, S., Hudry, B., Saadaoui, M., and Graba, Y. (2009) Classification of sequence signatures: a guide to Hox protein function. Bioessays 31, 500 –511 Le Roux, I., Joliot, A. H., Bloch-Gallego, E., Prochiantz, A., and Volovitch, M. (1993) Neurotrophic activity of the Antennapedia homeodomain depends on its specific DNA-binding properties. Proc. Natl. Acad. Sci. U. S. A. 90, 9120 –9124 Balayssac, S., Burlina, F., Convert, O., Bolbach, G., Chassaing, G., and Lequin, O. (2006) Comparison of penetratin and other homeodomain-derived cell-penetrating peptides: interaction in a membrane-mimicking environment and cellular uptake efficiency. Biochemistry 45, 1408 –1420 Blaber, M., DiSalvo, J., and Thomas, K. A. (1996) X-ray crystal structure of human acidic fibroblast growth factor. Biochemistry 35, 2086 –2094
The FASEB Journal 䡠 www.fasebj.org
Received for publication July 26, 2012. Accepted for publication October 1, 2012.
BECHARA ET AL.