Bright, M., P.C. Dworschak & M. Stachowitsch (Eds.) 2002: The Vienna School of Marine Biology: A Tribute to Jörg Ott. Facultas Universitätsverlag, Wien: 137-159
Phoenix from the Ashes? Repetitive Mass Mortality and the Biogeology of Southern Arabian Gulf (United Arab Emirates) Coral Systems Bernhard Riegl National Coral Reef Institute, Oceanographic Center, Nova Southeastern University, 8000 N. Ocean Drive, Dania 33004, FL, USA, and Institut für Geologie und Paläontologie, Karl-Franzens-Universität Graz, Heinrichstrasse 26, 8010 Graz, Austria. E-mail:
[email protected] Keywords: Biogeology, coral, sea surface temperature anomaly, mass mortality, Arabian Gulf
Abstract. Biogeological dynamics consist of a combination of community-ecological responses based on individual-species physiological tolerance limits to a dominant environmental stressor and their consequences on carbonate sedimentological properties. This interplay was investigated in coral carpet systems (biostromal coral frameworks) in the southern Arabian Gulf in Abu Dhabi and Dubai (UAE). Five coral assemblages occur in the area: (A) low coral cover, widely spaced Porites lutea (B) high coral cover (biostromal framework), dense Acropora spp. (C) low coral cover, widely spaced faviids (D) low coral cover, individual Siderastrea savignyana only (E) high coral cover (biostromal framework), dense Porites harrisoni. Coral community dynamics and the potential for the accretion of reefal limestones was found to be controlled by recurring sea-surface temperature anomalies. The effects of these heating or cooling events are mass mortality of all Acropora species. COADS data and time-series of AVHRR satellite data suggest potential recurrence periods of 11-17 years. Extreme positive SST anomalies only appeared after 1986. These events result in large areas of dead framework with only interspersed live corals (the non-Acropora members in the assemblages). Dead frameworks are intensely biocorroded and break down rapidly. Thus the next generation of framebuilders has to settle on the same substratum as the previous
138
Riegl
generation and no reef-structure, which would require several generations of corals growing on top of each other, is built. However, since only limited space is available for coral settlement (most of the seabed being sandy), these recurrent mass mortalities of a dominant space-competitor allow competitively weak species to persist in the system and thus maintain diversity. Strong recruitment pulses from surviving pockets of Acropora ensure the survival of this community-component in time.
Problem The phoenix is "a legendary bird …living five or six centuries in the Arabian desert, being consumed in fire by its own act, and rising in youthful freshness from its own ashes and often regarded as an emblem of immortality or of the resurrection…" (Gove, 1964). The phoenix not only stands for resurrection, its image is also used as "…a paragon of excellence or beauty…" (Gove, 1964) and as such is in more than one way a befitting analogy for the biogeological dynamics of Arabian Gulf coral systems. It is accepted fact that sea surface temperature anomalies and associated mortality events can cause widespread perturbations in coral systems (Glynn, 1996; Wellington et al., 2001). It is also well known that repeated severe perturbations will distance any ecological system further from a climax stage (Connell, 1978; Sousa, 1984). Coral reefs have long been viewed as complicated climax or nearclimax systems whose persistence depended on the environment remaining within relatively narrow constraints within the reef’s physiological optimum. Over the past few decades, with perturbations in reefal systems being common-place and well studied (Porter & Meier, 1992; Done, 1992; Glynn, 1996; Connell, 1997; Mumby et al., 2001), the dynamic nature of coral-associated systems has become apparent. Also the associated diversity of organisms, the highest in any ecosystem except rainforests, has since been identified as linked to, and often dependent on, disturbances. Since coral-associated sedimentary systems have the potential to deposit and retain significant amounts of skeletal calcium carbonate, the health of the coral system leaves traces in the geological record; only if corals can live long enough, can a reefal or biostromal carbonate structure be built. Tight organismenvironment feedback has been identified (Longman, 1981; Riegl & Piller, 2000). High latitude coral systems march to a different tune than their counterparts in the tropics. Subtropical seas are characterized by harsher climactic conditions, generally with higher temperature, salinity and nutrient fluctuations. This is particularly true for the Arabian Gulf, which for hermatypic corals certainly represents
Mass mortality and biogeology of S Arabian Gulf coral systems
139
one of the most extreme environments (Coles, 1988; Sheppard & Sheppard, 1991, Coles & Fadlallah, 1991, Sheppard et al., 1992, Riegl, 1999). Nevertheless, a relatively high biodiversity of typical reef-associated taxa occurs, with 34-50 coral species, depending on taxonomic views and study area (Basson et al., 1977; Hodgson & Carpenter, 1994; Riegl, 1999; Veron, 2000), and over 100 fish species (Krupp & Almarri, 1996; Carpenter et al., 1997; Riegl, 2002). Temperature anomalies appear to be increasing in frequency over much of the tropics (Lough, 2000), and 1998 brought the most significant hitherto recorded world-wide coral-bleaching and mortality event (Wilkinson, 1998, 2000; Goreau et al., 2000). In the southwestern Arabian Gulf, however, no less than four major temperature anomalies (either positive or negative) with associated coral massmortality have been recorded (1964: Shinn, 1976; about 1981: Holt-Titgen, 1982; summer 1996: Riegl, 1998, 1999; summer 1998: Goreau et al., 2000, Wilkinson, 2000, Riegl, 2002). For a coral-associated system, these mass mortality events appear to occur at an astonishingly rapid recurrence period that, due to the generally slow growth rate of corals (1-10 cm per year, Bossher, 1992; Coles & Fadlallah, 1991; Heiss, 1994), would raise the question how such systems can persist. The question about persistence automatically translates into one about framebuilding – if corals die so often, how does this influence their ability to build a coherent carbonate structure? Despite the harshness of the environment (Kinsman, 1964; Sheppard et al., 1992; Coles & Fadlallah, 1991; Fadlallah et al., 1995) and apparently recurring mass mortality (Riegl, 1999, 2002), the Arabian Gulf nevertheless harbors important and diverse coral areas. This could only be explained by a surprising ability of these systems to overcome repeated crises – and an ability to rise like the phoenix from their ashes, rejuvenated and beautiful. The present study therefore explores 1) the climatic triggers for coral mass mortality in the Arabian Gulf 2) its effects on coral community structure 3) its effects on framework building 4) the mechanisms of persistence and regeneration.
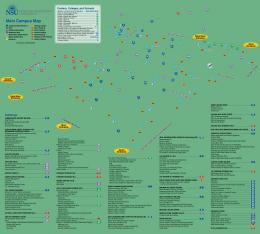
Material and Methods Study areas covered virtually the entire length of the coastlines of the Emirates of Abu Dhabi and Dubai (United Arab Emirates). This equates to about one fifth of the Arabian Gulf’s entire southern mainland coastline. The most closely studied area is situated in Dubai Emirate between Jebel Ali port and Ras Hasyan (Fig. 1). Coral areas in the region were studied annually between 1995 and 2001, although not every area was visited repeatedly. Coral areas were identified by a variety of
140
Riegl
Fig. 1: Study areas in the southern Arabian Gulf along the shorelines of Abu Dhabi and Dubai emirates (United Arab Emirates). The study areas are arranged into two geomorphologically distinct regions. The western study region consists of the coral areas at Muquaraq and Ruweis to the west and east of the headland formed by Jebel Dhannah. The Saddiyat coral area is in the channel between Abu Dhabi island and Saddiyat. The eastern study area stretches from Ras Hanyurah, where only few isolated corals are found, to Ras Ghanada, with big and well-developed framework carpets, via Ras Ghantoot and Ras Hasyan, the second dense and well-developed framework area, via Jebel Ali to Deira, directly off the town of Dubai.
methods ranging from side-scan-sonar, to diver grid-surveys, to image classification of aerial and satellite images (for further information see Riegl, 1999 and Riegl et al., 2001). Repeated quantitative sampling only took place in the Jebel Ali study area (1995, 1996, 1998, 1999), where twenty-six evenly distributed, equidistant sample points were selected within the known area of dense coral growth and two line transect sampling methods were utilized because of variable coral cover. Sampling in 1995/6 employed continuous intercept recording on 10 m line transects and line point intercept recording in one meter intervals on 50 m line transects (Loya, 1978; Weinberg, 1981; English et al., 1994). Details and rationale of the sampling
Mass mortality and biogeology of S Arabian Gulf coral systems
141
method can be found in Riegl (1999). After the 1996 coral mass mortality, the area was revisited and re-sampled in April 1998 (prior to the onset of the 1998 SST anomaly), October/November 1999 and July 2000 (no sampling). This time, only random transects were used in the same areas where sampling was performed in previous years (as recorded by d-GPS). The random sampling approach relied on overall statistical differences within the sampled assemblage rather than on repetitive measurements of the same colonies. Skeleton breakdown and renewed recruitment were documented. All coral recruits were identified to species and measured. Also areas that were known to have been affected by man-made disturbances (similar mortality due to dredging plumes in nearby Al Jazira close to Ras Ghantoot, Abu Dhabi) 10 years prior to the 1996 temperature anomaly were visited to obtain a time series of framework breakdown or regeneration processes. Data processing for classification and ordination used a square matrix of coral cover; data were fourth root transformed prior to the production of a similarity matrix. Agglomarative, hierarchical cluster analysis used the Bray-Curtis quantitative similarity coefficient and group average sorting (Warwick & Clarke, 1993) for 10-m transects. Although the Bray Curtis similarity coefficient is superior to Euclidian Distance as a measure (Bloom, 1981), the latter was nevertheless used for the 50-m transects since the clusters obtained with this algorithm were better interpretable from a biological viewpoint. Multidimensional scaling was used to provide an alternative to view the processes of change within assemblages (Clarke & Warwick, 1998). Coral taxonomy follows Wallace (1999) and Veron (2000). Acropora clathrata according to Wallace (1999) is now A. downingi. According to Veron (2000) both species names, A. downingi and A. clathrata, are valid, which is the opinion followed in this paper. Also, Porites compressa is renamed as Porites harrisoni (Veron, 2000). The taxonomy in this paper therefore differs from that used in a previous paper from the same study area (Riegl, 1999). Sea surface temperature (SST) was obtained by in situ measurements provided by the Dubai municipality (3 month records only) and satellite-derived data (monthly averages) were obtained from the website http://psbsgi1.nesdis. noaa.gov:8080/PSB/SST/climo_archive. These data go back to 1984. Further information was derived by AVHRR satellite imagery of the area provided by NOAA through various websites. Also an SST dataset from 1926 to 1992 was obtained from the COADS dataset (http://ingrid.ldeo.columbia.edu/sources) for the southeastern Arabian Gulf. For the estimation of spatial cover of coral frameworks, maps produced by Riegl et al. (2001) were imported into a ZEISS KS 400 3.0 image analysis program which allowed the determination of total space covered by corals in the surveyed
142
Riegl
area, versus areas covered by bare sand and seagrass. Thus the total area of framework formation potential could be calculated both in km² and percentage of total mapped area. This calculation ignored assemblage-specific space cover by corals within the used categories since it was only interested in the total area covered by coral growth (total square meterage covered by dense Acropora assemblage, dense Porites harrisoni assemblage, sparse Porites lutea assemblage, etc.). The same transects as used for coral assemblage analysis were used for disease counts. If any part of an intercepted colony was diseased, even if not directly under the transect line, it was recorded. This allowed detection of areas with high disease frequency and quantification of affected colonies in the population as well as species-specific frequency.
Results 1. Temperature anomalies Negative temperature anomalies occur in winter, caused by the Shamal, a cold north wind falling into the Gulf basin from the Iran highlands, that can cause severe chilling. Shinn (1976) attributed massive coral die-back in the southern Gulf (Qatar) to a severe Shamal, which caused surface cooling to 4 ºC and to 14.1 ºC. at 18 m depth. Also Coles & Fadlallah (1991) recorded coral death in Tarut Bay, Saudi Arabia, in 1983 due to extreme cold temperatures. The storm seas associated with strong Shamals can be significant (6 m wave height caused by 65 km·h-1 winds: Shinn, 1976; Sheppard et al., 1992) and cause good vertical mixing of the water column, thus allowing the drop in temperature to also affect deeper biota. Associated with Shamals is also a negative surge (up to -3 m) produced at the northern end of the Gulf, which can cause abnormally low tides (Fadlallah et al., 1995). Downing (1985) and Fadlallah et al. (1993) documented the mechanism in Kuwait and Tarut Bay, Saudi Arabia, and cooling by strong Shamals, often coupled with aerial exposure due to the extremely low tides, is a recognized factor shaping the existence of Gulf coral communities by causing repeated mortality (Shinn, 1976; Coles & Fadlallah, 1991; Fadlallah et al., 1995). Sustained positive temperature excursions that raised average SST by over 2 degrees for almost three months occurred in the southern Arabian Gulf in 1996 and 1998. In both years, the SST anomaly started in April and lasted until September (Riegl, 2002). June, July and August of both years showed a +2-2.5 °C anomaly, which is generally considered to be above the bleaching threshold. Maximum in situ measured temperatures in the study area were 35 °C, which is
Mass mortality and biogeology of S Arabian Gulf coral systems
143
about 2 degrees above the usually recorded maxima and thus supports the satellitederived data. The climatic factors associated with these positive anomalies are not well understood. Both years could have been influenced by the onset and senescence of the exceptionally strong 1997 El Niño Southern Oscillation that affected the Asian monsoon system and thus likely also the climate in the Arabian Gulf region. For the examination of past temperature anomalies likely to have caused mass mortality or at least bleaching, the COADS dataset was used. This dataset shows clearly that cold anomalies were more frequent than hot anomalies in the 20th century. Several events where temperature dropped to 14 °C below the century mean (26.7 °C) were recorded. It is likely, however, that several lethal events are not represented in the dataset because individual data points are averages from a relatively large area and a whole month. Extreme cold events are of short duration (24-72 h, Fadlallah et al., 1995) and therefore likely to be lost in the averaging process. The same may be true for warm anomalies, except for those of 1996 and 1998 which lasted several months. It appears that warm anomalies only started to become important in the later part of the century (after 1980). This would be consistent with warming trends observed by Lough (2000). Not every extreme excursion of the temperature curve automatically leads to a coral mass mortality and it has been suggested that at least another additional stressor (such Table 1: Quantitative biological characteristics of coral assemblages in the eastern study area between Jebel Ali and Ras Ghantoot. Dominant species are those that cover the most space within each assemblage (i.e. ecologically and visually dominant). Not all assemblages were sampled by both transect methods (n.s. = not sampled). Dominant typology refers to the growth-form type found in most constituent corals of the respective assemblage. "Big massive" means that corals usually attained sizes up to and over 50-100 cm diameter, "small massive" means that corals were usually less than 10 cm in diameter. Values are means and standard deviation of percentages of total transect (sample) area covered by each assemblage (all species). assemblage dominant species code (alive) in 1996
A B C D E
Porites lutea Acropora clathrata and A. downingi Favia spp. Siderastrea savignyana Porites harrisoni
living coral cover of substratum, all species (10m transects) 36±15 62±24
living coral cover of substratum, all species (50 m transects) 21±11 54±22
dominant typology of corals
16±4 n.s.
n.s. 14±7
big massive tabular branching small massive small massive
n.s.
51±2
columnar
144
Riegl
as exposure to air or sedimentation) may be needed to trigger mass mortality (Fadlallah et al., 1995; Riegl, 1999). Records for previous mass mortality in the study area exist in the literature. Holt Titgen (1982) speculated on timing of a mass mortality near Jebel Ali in 1981, Shinn (1976) suggested that a coral mass mortality in Qatar (and therefore likely also at least in the western study area in the UAE) could have been caused by an unusually strong Shamal in 1964. Other reports, particularly from Saudi Arabia, may be too far away to have bearings on the present study. SST records from the COADS dataset support these findings. Therefore, if only the most extreme events are considered, supported by literature reports on mass mortality, then possible years for previous mass mortalities could be: 1936 (14 °C below century mean), 1949 (14 °C below century mean), 1964 (Shinn, 1976, but no extreme anomaly in COADS dataset), 1981 (Holt Titgen 1982, but no extreme excursion in data set), 1985 (14 °C below century mean), 1996 (9 °C above century mean), and 1998 (9 °C above century mean).
2. The structure of coral assemblages and biological effects of temperature anomalies Coral assemblage structure was quantitatively investigated at Jebel Ali, Ras Hasyan and Ras Ghantoot. It was qualitatively assessed at Muqaraq, Ruweis, Saddiyat, Al Taweelah, Ras Ghanada, and Deira. Prior to the 1996 mass mortality, five well separable coral assemblages of highly variable substratum coverage were differentiated by both 10 m and 50 m transects (Table 1). A: large, widely spaced Porites lutea and other Porites with other, mainly massive, species. This coral assemblage had the highest spatial extension and did not suffer during either the 1996 or the 1998 SST anomaly (Tables 1, 2). B: mainly tabular colonies of Acropora clathrata and A. downingi and six other, generally subordinate Acropora species with high (40-90%) coral cover of available substratum. Frequent overtopping of competitively subordinate massive corals, like Porites spp., Cyphastrea spp. and Platygyra spp. Towards its periphery, cover by other Acropora species (such as A. pharaonis) and massive corals of the genera Porites, Cyphastrea, Favia and Platygyra increased. Heavy mortality in the 1996 mass mortality event (Fig. 2, A-B, Tables 1, 2), not in 1998. All Acropora removed. C: widely spaced patches of faviid colonies that were not affected either in 1996 or 1998. D: widely spaced Siderastrea savignyana colonies that were also not affected either in 1996 or 1998.
Mass mortality and biogeology of S Arabian Gulf coral systems
145
Table 2: Characteristics of coral assemblages in the western study area between Jebel Ali and Ras Ghantoot after the 1996 coral mortality event. Coral cover refers to the percentage of transect samples that were covered by live corals or unbroken skeletons (= standing dead) without any living tissues. Values are means and standard deviation of percentages. Employed tests were one-way ANOVA with Student-Neuman-Keuls post-hoc tests, or ttest. n.a. = not applicable.
% total coral cover of substratum (live and standing dead)
1996
1998
1999
significance
post hoc difference
90±12
83±11
81±6
F = 2.05 p > 0.05
n.a.
26±6
22±10
F = 144,3 p < 0.01
1996-1998 1996-1999
0
0
n.a.
n.a.
% living coral cover of substratum (no standing dead included) 90±12 % contribution of live Acropora to total cover
80±22
E: densely spaced (80% coral cover) columnar Porites harrisoni colonies intermingled with massive colonies, mainly faviids (Favia sp., Platygyra sp.) with patchy distribution. Low mortality in 1996 and 1998. At Muqaraq in November 1999, the coral area was small and consisted mainly of assemblages C and D, with only a small (a few dozen square meters) area of assemblage A, which by that time had already completely broken down and only consisted of rubble. At Ruweis in November 1997, all four assemblages were present and covered a large area. All had suffered significant mortality in the 1996 SST anomaly. At Saddiayat in July 2000, only Porites-dominated patch-reefs were investigated. The Acropora component had been totally removed, either in 1996 or 1998. Near Al Taweelah, in October 2001, mainly assemblages A, C, and D occurred with only small and isolated patches of assemblage B. The densest areas with several square kilometres of assemblage B were found at Ras Ghanada and Ras Hasyan (between 1995 and 2001). These areas also had all other assemblages. In both areas, assemblage B had lost all Acropora in 1996. Smaller, but nevertheless extensive areas of assemblage B were found near Ras Ghantoot (between 1995 and 2001) and at Deira in April 1998. In both areas, assemblages A and C were also encountered. The Al Jazira population near Ras Ghantoot and the Deira population only suffered partial Acropora mortality in 1996 and 1998. The SST anomaly in 1996 affected primarily Acropora species in all sample areas and had negligible effects on other taxa in the eastern study area between Al Taweelah and Deira, but also caused significant mortality among other taxa in the
146
Riegl
western area (Muqaraq and Ruweis). The total coral fauna in the eastern area was reduced from 34 species to 27 species (all Acropora removed, fauna check-list in Riegl, 1999); changes in the western area were not recorded because no exhaustive pre-mortality surveys were conducted. However, the effect is assumed to be similar. Since Acropora had the highest framebuilding capacity (high population density, interlocking and rapid skeletal growth), the living component of virtually the entire framework-producing area (with the exception of the dense Porites harrisoni assemblage) was lost in this mass mortality event. Dense Porites harrisoni frameworks did not suffer significantly in the eastern study area, but suffered some mortality in the western study area at Ruweis. Maps of the eastern study area between Jebel Ali and Ras Hasyan prior to the mortality event showed 7.9 km² with framebuilding potential of the total investigated 37.7 km². After the 1996 SST anomaly and associated Acropora mass mortality, this was reduced to 0.1 km² – a loss of 98.7% (Fig. 3). The 1998 SST anomaly had only minor effects on the assemblage and triggered neither mass bleaching nor strong mortality. This is most likely because all susceptible species had already been removed from the coral assemblage. The SST anomaly was as strong as that of 1996. The other large coral area in the eastern study area, off Ras Ghanada, showed the same pattern of mortality, although the area of frameworks was not quantified. It is not known whether Acropora mass mortality occurred in 1996 or 1998. Fig. 2: Biogeological dynamics in Arabian Gulf coral communities: (A) in 1995, healthy Acropora (in the foreground A. arabensis, in the background A. downingi tables) dominate the coral system (in the right background an A. downingi table overshadows a Porites lutea). Eastern study area, Ras Hasyan (B) In 1996, all Acropora died due to a positive SST anomaly, most non-acroporid taxa survived (seen here are Platygyra lamellina, Cyphastrea microphthalma, and Porites lichen). Eastern study area, Ras Hasyan. (C) In 2001, five years after the mass mortality, even slow-growing species, like this Porites nodifera, reach above the former Acropora canopy. Surrounding the Porites are dead Acropora tables. Eastern study area, Ras Ghanada. (D) After mortality, the colonies remain standing for a few years, before they start to break down. Western study area, Ruweis, in November 1997. (E) Five years after mortality, in Nov. 2001, Acropora skeletons start breaking down and form rubble beds among the surviving and thriving Porites. Eastern study area, Ras Ghanada. (F) Ten to fifteen years after death, most Acropora skeletons are turned into rubble. Jebel Ali in 1995, corals were killed between 1980 and 1985 by dredging and nearby construction. (G) A one- or two-year old-recruit in the eastern study area at Ras Hasyan in October 1999. This colony would have settled either in 1997 or 1998, suggesting that some areas survived and may have even remained fecund during the 1998 SST anomaly. (H) onward on the path to regeneration. A 3-4-years-old recruit in November 2001 in the eastern study area at Ras Ghanada.
Mass mortality and biogeology of S Arabian Gulf coral systems
147
148
Riegl
3. Framework breakdown After the Acropora mass mortality of 1996 (in April 1998), most Acropora skeletons were still standing and branches were covered by fine algal turf without heavy overgrowth. Erosion of surficial structures by grazing echinoids (particularly Echinometra mathei) was evident (Figs 2, 4). Coral skeletons were still rigid and not easily broken. In November 1999, branches were encrusted by layers of oysters (Chama sp., Spondylus sp.) and coralline algae. Boring by clionid sponges rendered many branches brittle. In 1999, many Acropora tables had started to degenerate into stumps surrounded by branch fragments. In a coral area west of Ras Ghantoot that had been killed 10 years previously (1985/6), virtually no intact Acropora tables remained (Fig. 2 E, F). In 2001, breakdown continued and many tables had obtained a dense overgrowth by coralline algae and the green alga Lobophora variegata (Fig. 2 C). The mechanism of framework breakdown due to bioerosion and physical forces was observed in all study areas but recorded in detail in the vicinity of the Al Jazira channel near Ras Ghantoot; this area had suffered mortality and delayed or no recruitment due to smothering by a dredging spoil plume between 1980 and 1985 (personal communication by the dredging engineer). Thus, in comparing the areas, a clear sequence of framework breakdown could be established. In Al Jazira, ten to fifteen years after coral mortality, all tables had broken down and the previous framework now consisted only of well-spaced stumps. This finding suggests that about 15 years after a mass mortality most of the Acropora framework is removed. The breakdown sequence was as follows (Fig. 4): In the first years, from 1997 to 1998, only algal turfs and surficial erosion due to echinoids was observed. The settlement of a single, shortlived layer of Chama aspera also occurred In 1999, 3 years after mortality, overgrowth by coralline algae, bivalves (mainly Spondylus) and weakening of skeletons due to clionid sponges and boring bivalves was observed. In 2001, five years after death, heavy overgrowth by bivalves and corallinacean algae fused the open latticework of branches on most tables to solid plates. Heavy internal biocorrosion by clionid sponges and boring bivalves had significantly weakened the skeletons. The framework was brittle and started to break under its own weight.
4. Recruitment and regeneration In October 1999, recruits near Ras Ghantoot ranged in size from 10-70 cm in greatest diameter (average 34 ± 15 cm), which would suggest a growth rate between 5 and 10 cm per year since they had had a maximum of 14 years to grow
Mass mortality and biogeology of S Arabian Gulf coral systems
149
Fig. 3: Map of coral assemblage distribution in the western study area between Ras Hasyan and Jebel Ali, Dubai, UAE. The striped area was covered by dense Acropora growth in 1995 and only by standing dead skeletons after summer 1996. The small dark grey area was covered by a columnar Porites harrisoni assemblage, the light grey area by the massive Porites lutea-dominated community. The white area is mainly sand, but includes patches of Siderastrea and faviid-dominated, sparse coral associations. Seagrass and algae beds are indicated with a leaf texture.
(earliest possible settlement in 1985). This is consistent with measurements by Coles & Fadlallah (1991) from Saudi Arabia. In October 1999, recruits in the Ras Hasyan and Jebel Ali area had an average diameter of 7.6 ± 3.5 cm (n = 53). This suggests an annual growth rate of about 10 cm per year (see above and Coles & Fadlallah 1991) since most recruits would have settled after summer 1998, when
150
Riegl
conditions once again allowed survival. A few big recruits (diameter 25cm, n = 3) probably already settled in 1997 and survived the 1998 SST anomaly, but were overlooked in the previous years. Recruits near Al Taweelah in 2001 were between 20 and 40 cm in diameter, which suggested settlement in 1998. Most grew on the caprock (original rocky attachment substratum), with only a few settling on dead corals.
5. Effects on coral diseases Black Band Disease (BBD) was the most common disease on branching corals during summer 1995 but disappeared in winter 1995/6 and after the mass mortality in 1996. Prior to the coral mass mortality, some areas had 25% infection rates on A. clathrata and A. downingi. White Band Disease (WBD) was rare both prior to and after the mass mortality. In 1998/9 it was still found on the same species except Acropora. Yellow Band Disease (YBD) was widespread from 1995 through 1999 both in summer and winter. In 1995/6, coral diseases were most frequent in areas of high coral coverage, particularly in Acropora-dominated areas. Space cover and percentage of diseased colonies was significantly positively correlated (Kendall’s Tau = 0.73, p = 0.029). After the 1996 SST anomaly and Acropora mass mortality, any density effects in coral diseases were lost. Coral diseases were seasonal prior to the mass mortality. In areas where BBD dominated in summer (1995), there was a shift towards a higher frequency of YBD in winter (1995/6). After 1996, seasonality (14±5% of corals, mainly Acropora, affected in summer, in winter 7±6%, mainly massives) was lost and infection remained on winter-levels (6±5%, only massives infected).
Discussion Coral ecologists have long come to realize that coral reefs are not stable systems and indeed shaped very much by disturbances (Done, 1992; Tanner & Hughes, 1996; Connell, 1997; Done, 1999). Studies such as the present one show how dynamic coral systems can be and how much disturbance they are able to withstand. This study provides evidence that temperature-related mass mortality in the Arabian Gulf is a recurrent phenomenon. These mortalities can either be caused by negative (Shinn, 1976; Coles & Fadlallah, 1991) or positive (Riegl, 1999; Goreau et al., 2000) temperature anomalies. In Abu Dhabi and Dubai, the latest important coral mass mortality was triggered by the 1996 positive SST anomaly,
Mass mortality and biogeology of S Arabian Gulf coral systems
151
Fig. 4: Schematic illustration of the processes involved in Acropora bioerosion as observed in all study areas in the United Arab Emirates.
152
Riegl
with negligible mortality in 1998, a year of similarly elevated temperatures. Since the 1996 SST anomaly affected the entire southern Gulf, some of the mortality attributed in the literature to the 1998 SST anomaly (p. ex. Goreau et al., 2000) may well have already happened in 1996. The 1996 mortality event is the first documented large-scale coral kill by a positive SST anomaly in the Arabian Gulf, while kills by negative SST anomalies have been repeatedly reported previously (Shinn, 1976; Coles & Fadlallah, 1991; Fadlallah et al., 1995). This is also supported by the COADS dataset, which shows cold anomalies as having recurred frequently in the 20th century, wheeas hot anomalies have not. Effects of the 1996 SST anomaly were detectable on several levels. On a coral assemblage level, a total shift of dominance away from branching Acropora species, which all died, to the surviving massive taxa (of which Porites was the most common) occurred (Riegl, 1999). This corresponds well with data from other areas in the Indo-Pacific (Fisk & Done, 1985; Brown & Suharsono, 1990; Gleason, 1993; Marshall & Baird, 2000), where Acroporidae were found to be among the most sensitive taxa to bleaching stress, and Poritidae were less and Cyphastrea the least susceptible. Some degree of adaptation to these apparently recurring mass mortalities may be a mechanism of community maintenance used by such high-latitude assemblages: the Acropora species that dominate in "good" years are competitively far superior to most other corals (Thomason & Brown, 1986) and would, if left unchecked, completely outcompete them. Since only limited space is available on the generally sandy seafloor, this mechanism would, probably in as little as a century, lead to a near-complete marginalisation of all slower-growing corals. This was already clearly visible in "ripe" Acropora clathrata and A. downingi stands in 1995/6, after about 10-15 years of undisturbed growth, where most faviids and poritids were overshadowed by the tabular Acropora, which formed an increasingly closed canopy (Riegl, 1999). After Acropora mortality, the slower-growing faviids and poritids were able to once again grow out from under the shadows of the slowly disintegrating Acropora tables (Fig. 5). The 1998 bleaching event was considered by many as the most important bleaching event ever documented, largely because of the magnitude of coral destruction and the wide geographical area affected (Goreau et al., 1999). However, the Arabian Gulf experienced comparable bleaching already in 1996 and not in 1998 (Wilkinson, 1998; Riegl, 1999, 2002). The data set presented here, as well as reports from the literature (Shinn, 1976; Holt-Titgen, 1986) suggest that such mortalities may indeed be recurring events, at least at a local scale. While undisputedly destructive and startling in their severity, they have to date not hindered the existence of rich coral areas even in extreme latitudes. The fact that mainly Acropora species were affected by the here-described mass mortality events, with less damage to other taxa, has parallels elsewhere. While
Mass mortality and biogeology of S Arabian Gulf coral systems
153
this preferential mortality is extremely well developed in the Arabian Gulf, Acropora species also suffered preferentially in the Indo-Pacific during cyclones, Acanthaster planci (crown-of-thorns seastar) outbreaks, red tides (Pearson, 1981) and bleaching events (Coles & Fadlallah, 1991; Gleason, 1993; Wilkinson, 1998; Marshall & Baird, 2000) as well as in the Atlantic reefal province due to disease outbreaks and destruction by hurricanes (Aronson & Precht, 1997). While in particular Atlantic Acropora species have suffered severe restrictions in frequency in the wake of mass mortality, Arabian Gulf Acropora appear to be able to recover from such major disturbances. It is interesting to note that not all Acropora in the wider study area died. Surviving pockets of healthy Acropora existed in 1999 at Deira and at Ras Ghantoot (near Al Jazira). This also corresponds well to data from the literature (Marshall & Baird, 2000). It was found that bleaching severity could differ between sites and that acclimatization due to a more severe bleaching history could help corals survive. In the study area, the surviving Acropora were close to major channels (Al Jazira population close to Al Jazira channel, a major channel leading to enclosed basins inland. Water with increased temperature and salinity drains as density flows (i.e. distinct water masses which generally mix little with the surrounding water; Lang et al., 1988; Hickey et al., 2000) along the bottom of the channel and is likely to repeatedly impact the nearby coral population) and ports (Deira population adjacent to Port Rashid, a major industrial port). These populations are more likely to have repeatedly experienced severe environmental stresses than the corals in more open ocean settings in either the eastern or western study area and may therefore be better acclimatized to higher-than-normal temperatures. Since Acropora may indeed "come and go" on a temporal scale, other system components which appear to depend on high density of a particular coral species, like coral diseases or certain fish assemblages, could also be expected to change after such a significant mass mortality event. Corresponding to the loss of Acropora, coral diseases that preferentially occurred on Acropora (like BBD) disappeared from the area (western study area Ras Ghantoot to Jebel Ali). Prior to the 1996 coral mass mortality, high permanent levels of coral diseases with preferential distribution in dense coral areas were encountered, a situation also reported from the northern Arabian Sea (Coles, 1994, 1997; Coles & Seapy, 1998). The 1996 Arabian Gulf mass mortality provides evidence that the frequency of diseases could in some cases be regulated by the availability of preferred "target species". Coral diseases acted density dependent and appeared to have evolved to target the most common species. When all Acropora died in 1996, at least one disease (BBD) had virtually disappeared, and since fewer corals were left overall, coral diseases decreased in frequency (Riegl, 2002).
154
Riegl
From a framebuilding perspective, the relatively rapid breakdown of coral skeletons (Figs 6, 10) demonstrates the importance of bioerosion and biocorrosion in framework breakdown (Glynn, 1988, 1990, 1997; Hibino & van Woesik, 2000). While on the outside of the skeletons significant bioaccretion occurred (in particular settlement of bivalves and polychaetes), the mechanical stability of the whole structure was compromised by clionid sponges and mytilid mussels boring the inside of the branches and the stems of the tables (Fig. 6). Similar observations are reported in the literature (Tunnicliffe, 1979; Highsmith, 1981; Scott & Risk, 1988; Hassan, 1998). Since virtually the entire framebuilding coral population died at the same time, skeleton breakdown could proceed at the same pace in the whole area, probably destroying most of the Acropora framework over the entire area and leading to removal of the fragments by waves and currents. Had only individual colonies died and broken down, their fragments would have remained trapped inbetween the living corals, which could have led to reef buildup. The resultant structure could have been a fringing reef. One possible generalization from this study is that catastrophic mortality of entire coral populations can have negative repercussions on framebuilding potential, while death of individual colonies need not. Note, however, that different processes could be taking place in other parts of the Arabian Gulf, in particular the offshore islands, which are not included in the present study. Active sexual recruitment into the area was observed already two years after the mass mortality. The first few recruits could even have arrived in the first year after the mortality. Seeding populations, although also undoubtedly stressed by the thermal anomaly, nevertheless apparently completed their reproductive cycle. Fadlallah et al. (1992) and Fadlallah (1996) observed Acropora spp. to spawn even after a major negative temperature anomaly that caused widespread coral mortality in Saudi Arabia. Although disturbances and stress are known to reduce coral fecundity (Szmant & Gassman, 1990; Hoegh-Guldberg, 1999) it appears that surviving Arabian Gulf corals were sufficiently fecund to reproduce and thus to replenish coral losses after such major stress events. This has important implications for the temporal persistence of the coral systems. The apparent different effects of the disturbances on competitively inferior species (poritids and faviids, Fig. 5) and the dominant competitor (Acropora) seems to support concepts for the maintenance of diversity in ecological communities like the intermediate disturbance hypothesis and the concept of compensatory mortality (Connell, 1978; Sousa, 1984). These hypotheses assume consistent asymmetrical outcomes in competitive encounters, with one of a pair of competitors always winning over the other (Sousa, 1984). For the weaker party to persist in the system, the stronger needs to be regularly, or preferentially, stunted (see also
Mass mortality and biogeology of S Arabian Gulf coral systems
155
Fig. 5: Model for framework destruction by Riegl (2001) as driven by repeated mass mortality or stunting of a dominant framebuilding genus (or guild), in this case Acropora. 1) the system is at maximum distance from the last disturbance. High space coverage by the dominant competitor (in this case Acropora) leads to high coral cover of available substratum and production of an interlocking coral framework. The dominant competitor outcompetes most other species, with negative effects on diversity. 2) A mass mortality removes the dominant competitor. Since the entire (or the largest part) of the population is wiped out, no larval reservoir is available for rapid recolonization. The coral framework is bioeroded and the fragments transported away. By the time sufficient recruits are available, the framework is already gone and the recruits have to settle on the same substratum as the previous generation. Therefore, no lasting framework is built.
Baird & Hughes, 2000). The conditions required by these hypotheses are by and large met in Arabian Gulf coral assemblages (Acropora was winning over all other species and preempted most space; competitively inferior species start to expand their space coverage after disturbance). The observed disturbing factors were coral diseases and SST anomalies, which both effected compensatory mortality in the dominant competitor. The evidence collected in this paper suggests that, like the legendary phoenix, Arabian Gulf coral assemblages are capable of resurrecting from the ashes of the temperature-mediated mass mortalities. The time required for the dominant competitor to recover allows the system's diversity to persist, if not to slowly increase, and thus renews the phoenix's beauty.
156
Riegl
Acknowledgements This paper is a result of Austrian Science Foundation (FWF) project P-13165-GEO but was additionally assisted by many individuals and organisations. Assistance in the field was rendered by (in alphabetic order) J. Ardon , D. Buerger, M. de Vries, M. Greiss, J. Korrubel, C. Martin, F. Sommer, J. Wijsman and many others who helped on boats, on shore and by granting permits. Research was encouraged and partly sponsored by the Dubai Municipality and various Abu Dhabi agencies. Special thanks go to H.K. al-Shaer, M.A. Hassan Deshgooni and S. Mustafa of the Dubai Municipality. Martin Mid East Ltd., Delft Hydraulics and the Abu Dhabi Water and Electricity Agency provided access to documentation, maps and logistic support in the field. W.E. Piller’s thoughtful input in many discussions is gratefully acknowledged.
References Aronson, R.B. & W.F. Precht, 1997: Stasis, biological disturbance, and community structure of a Holocene reef. Palaeobiology, 23(3): 326-346. Aronson, R.B., W.F. Precht, I.G. Macintyre & T.J.T. Murdoch, 2000: Coral bleach-out in Belize. Nature, 405: 36. Baird, A.H. & T.P. Hughes, 2000: Competitive dominance by tabular corals: an experimental analysis of recruitment and survival of understorey assemblages. J. Exp. Mar. Biol. Ecol., 251: 117-132. Basson, P.W., J.E. Burchard, J.T. Hardy & A.R.G. Price. 1977: Biotopes of the western Arabian Gulf. ARAMCO Saudi Arabia, Dahran; 284 pp. Bloom, S.A., 1981: Similarity indices in community studies: potential pitfalls. Mar. Ecol. Progr. Ser., 5: 125-128. Bosscher, H., 1992: Growth potential of coral reefs and carbonate platforms. Academisch Proefschrift, Vrije Universiteit Amsterdam: 1-157. Brown, B.E. & Suharsono, 1990: Damage and recovery of coral reefs affected by El Nino related seawater warming in the Thousand Islands, Indonesia. Coral Reefs, 8: 163-170. Carpenter, K.E., F. Krupp, D.A. Jones & U. Zajonz. 1997: Living marine resources of Kuwait, eastern Saudi Arabia, Bahrain, Qatar, and the United Arab Emirates. FAO Rome; 293 pp. Clarke, K.R. & R.M. Warwick, 1998: Quantifying structural redundancy in ecological communities. Oecologia, 113: 278-289. Coles, S.L., 1994: Extensive coral disease outbreak at Fahl Island, Gulf of Oman, Indian Ocean. Coral Reefs, 13: 242. Coles, S.L., 1997: Reef corals occurring in a highly fluctuating temperature environment at Fahl Island, Gulf of Oman (Indian Ocean). Coral Reefs, 16: 269-272. Coles, S.L. & Y.H. Fadlallah, 1991: Reef coral survival and mortality at low temperatures in the Arabian Gulf : new species-specific lower temperature limits. Coral Reefs, 9: 231-237.
Mass mortality and biogeology of S Arabian Gulf coral systems
157
Coles, S.L. & D.G. Seapy, 1998: Ultra-violet absorbing compounds and tumorous growths on acroporid corals from Bandar Khayran, Gulf of Oman, Indian Ocean. Coral Reefs, 17: 195-198. Connell, J.H., 1978: Diversity in tropical rain forests and coral reefs. Science, 199: 1302-1310. Connell, J.H., 1997: Disturbance and recovery of coral assemblages. Coral Reefs, 16, Suppl: S101-S113. Done, T.J., 1992: Constancy and change in some Great Barrier Reef coral communities: 1980-1990. Amer. Zool., 32: 655-662. Done, T.J., 1999: Coral community adaptability to environmental change at the scales of regions, reefs and reef zones. Amer. Zool., 39: 66-79. Downing, N., 1985: Coral reef communities in an extreme environment: the northwest Arabian Gulf. Proc 5th Int. Coral Reef Congr., 6: 343-348. English S., C. Wilkinson & V. Baker 1994: Survey manual for tropical marine resources. Aust. Inst. Mar. Sci., Townsville; 386 pp Fadlallah, Y.H., 1996; Synchronous spawning of Acropora clathrata coral colonies from the western Arabian Gulf (Saudi Arabia). Bull. Mar. Sci., 59: 209-216. Fadlallah, Y.H., K.W. Allen & R.A. Estudillo, 1995. Mortality of shallow reef corals in the western Arabian Gulf following aerial exposure in winter. Coral Reefs, 14: 99-107. Fadlallah, Y.H., R.T. Lindo & D.J. Lennon, 1992: Annual synchronous spawning event in Acropora species for the western Arabian Gulf. Proc 7th Int. Coral Reef Symp., 1: 501. Fisk, D.A. & T.J. Done, 1985: Taxonomic and bathymetric patterns of bleaching in corals, Myrmidon Reef (Queensland). Proc. 5th Int. Coral Reef Congr., 6: 149-154. Gleason, M.G., 1993: Effects of disturbance on coral communities: bleaching in Moorea, French Polynesia. Coral Reefs, 12: 193-201. Glynn, P.W., 1988: El Nino warming, coral mortality and reef framework destruction by echinoid bioerosion in the Eastern Pacific. Galaxea, 7: 129-160. Glynn, P. W., 1990: Coral mortality and disturbances to coral reefs in the tropical eastern Pacific. in: Glynn, P.W. (Ed.) Global ecological consequences of the 1982-83 El NinoSouthern Oscillation. Elsevier Oceanogr. Ser., 52, 55-126. Glynn, P. W., 1996: Coral reef bleaching: facts, hypotheses and implications. Global Change Biol., 2: 495-509. Glynn, P. W., 1997: Bioerosion and coral reef growth. In: C. Birkeland (Ed.), Life and death of coral reefs. Chapman and Hall, New York, 68-95, Goreau, T.J., T. McClannahan, R. Hayes, & A. Strong, 2000: Conservation of coral reefs after the 1998 global bleaching event. Conservation Biology, 14(1): 5-15. Gove, P.B., (Ed.) 1964: Webster’s third new international dictionary of the English language. G. & C. Merriam Company Publishers, Springfield, Mass.; 1699 pp. Hassan, M., 1998: Modification of carbonate substrata by bioerosion and bioaccretion on coral reefs of the Red Sea. Shaker Verlag, Aachen; 126 pp. Heiss, G.A., 1994: Coral reefs in the Red Sea: growth, production, and stable isotopes. Geomar Report, 32: 1-141.
158
Riegl
Hibino, K. & R. van Woesik, 2000: Spatial differences and seasonal changes of net carbonate accumulation on some coral reefs of the Ryukyu Islands, Japan. J. Exp. Mar. Biol. Ecol., 252: 1-14. Hickey, B.M., P. MacCready, E. Elliott, & N.B. Kachel, 2000: Dense saline plumes in Exuma Sound, Bahamas. J. Geophys. Res., 105: 11471-11488. Highsmith, R.C., 1981: Coral bioerosion at Eniwetak: agents and dynamics. Int. Rev. Ges. Hydrobiol., 66: 335-375. Hoegh-Guldberg, O., 1999: Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res., 50(8): 839-866. Holt Titgen, R. , 1982: The systematics and ecology of the decapods, and their zoogeographic relationship to the Arabian Gulf and the western Indian Ocean. Unpubl. Ph.D. Dissertation, Texas A and M University; 291 pp. Kinsman, D.J.J., 1964: Reef coral tolerance of high temperatures and salinities. Nature, London, 202: 1280-1282. Krupp, F. & M.A. Almarri, 1996: Fish and fish assemblages of the Jubail marine wildlife sanctuary. In: F. Krupp, A.H. Abuzinada, & I.A. Nader (Eds.), A marine wildlife sanctuary for the Arabian Gulf. Environmental research and conservation following the 1991 Gulf War oil spill. NCWCD, Riyadh and Senckenberg Research Institute, Frankfurt a.M.: 339-350. Lang, J.C., R.I. Wicklund & R.F. Dill, 1988: Depth- and habitat-related bleaching of zooxanthellate reef organisms near Lee Stocking Island, Exuma Cays, Bahamas. Proc. 6th Int. Coral Reef Symp., 3: 269-274. Longman, M.W., 1981: A process approach to recognizing facies of reef complexes. in: D.F. Toomey, (Ed.) European fossil reef models. SEPM Spec. Publ., 30: 9-40. Lough, J.M., 2000: 1997-98: Unprecedented thermal stress to coral reefs? Geophys. Res. Lett., 27(23): 3901-3904. Loya, Y., 1978: Plotless and transect methods. In: Stoddard, D.R., R.E. Johannes (Eds) Coral reefs: research methods. UNESCO, Paris: 197-217. Marshall, P.A. & A.H. Baird, 2000: Bleaching of corals on the Great Barrier Reef: differential susceptibilities among taxa. Coral Reefs, 19: 155-163. Pearson, R.G., 1981: Recovery and recolonization of coral reefs. Mar. Ecol. Prog. Ser., 4: 105-122. Porter, J.W. & O.W. Meier, 1992: Quantification of loss and change in Floridian reef coral populations. Am. Zool., 32: 625-640. Riegl, B., 1999: Corals in a non-reef setting in the southern Arabian Gulf (Dubai, UAE): fauna and community structure in response to recurrent mass mortality. Coral Reefs, 18(1): 63-73. Riegl, B., 2001: Inhibition of reef framework by frequent disturbance: examples from the Arabian Gulf, South Africa, and the Cayman Islands. Palaeogeogr. Palaeoclimatol. Palaeoecol., 175: 79-101. Riegl, B., 2002: Effects of the 1996 and 1998 positive sea surface temperature anomalies on corals, coral diseases and fish in the Arabian Gulf (Dubai, UAE). Mar. Biol., 140: 29-40.
Mass mortality and biogeology of S Arabian Gulf coral systems
159
Riegl, B., J.L. Korrubel & C. Martin, 2001: Mapping and monitoring of coral communities and their spatial patterns using a surface-based video method from a vessel. Bull. Mar. Sci., 68: 869-880. Riegl, B. & W.E. Piller, 2000: Reefs and coral carpets in the northern Red Sea as models for organism-environment feedback in coral communities and its reflection in growth fabrics. in: E. Insalaco, P.W. Skelton, & T.J. Palmer (Eds) Carbonate platform systems: components and interactions. Geol. Soc. (Lond.) Spec. Pub., 178: 71-88. Scott, P.J.B. & M.J. Risk, 1988: The effect of Lithophaga (Bivalvia: Mytilidae) boreholes on the strength of the coral Porites lobata. Coral Reefs, 7: 145-151. Sheppard, C.R.S. & A.L.S. Sheppard, 1992: Corals and coral communities of Arabia. Fauna of Saudi Arabia, 12: 3-170. Sheppard, C.R.S., P. Price & C. Roberts, 1992: Marine ecology of the Arabian region. Academic Press, London; 347 pp. Shinn, E.A., 1976: Coral reef recovery in Florida and the Persian Gulf. Environm. Geol., 1: 241-254. Sousa, W.P., 1984: The role of disturbance in natural communities. Ann. Rev. Ecol. Syst., 15: 353-391. Szmant, A.M. & N.J. Gassmann, 1990: The effects of prolonged “bleaching” on the tissue biomass and reproduction of the coral Montastrea annularis. Coral Reefs, 8: 217-224. Thomason, J.C. & B.E. Brown, 1986: The cnidom: an index of aggressive proficiency in scleractinian corals. Coral Reefs, 5: 93-101. Tanner, J.E. & T.P. Hughes, 1996: The role of history in community dynamics: a modelling approach. Ecology, 77(1): 108-117. Tunnicliffe, V., 1979: The role of boring sponges in coral fracture. In: C. Levy & S. BouryEsnault (Eds) Biologie des Spongiaires. CNRS, Paris, 291: 309-315. Veron, J.E.N., 2000: Corals of the world. 3 Volumes. Aust. Inst. Mar. Sci., Townsville; 463 pp., 429 pp., 490 pp. Wallace, C.C., 1999: Staghorn corals of the world. A revision of the coral genus Acropora (Scleractinia; Astrocoeniina; Acroporida) worldwide, with emphasis on morphology, phylogeny and biogeography. CSIRO Publishing; 421 pp. Warwick, R.M. & K.R. Clarke, 1993: A comparison of methods for analyzing changes in benthic community structure. J. Mar. Biol. Assoc. UK, 71: 225-244. Weinberg, S., 1981: A comparison of coral reef survey methods. Bijdr. Dierk., 51(2): 199218. Wellington, G.M., P.W. Glynn, A.E. Strong, S.A. Nauarrete, E. Wieters & D. Hubbard, 2001: Crisis on coral reefs linked to climate change. EOS, 82(1):1-7. Wilkinson, C., 1998: Status of the coral reefs of the world: 1998. Aust. Inst. Mar. Sci., Townsville; 184 pp. Wilkinson, C., 2000: Status of the coral reefs of the world: 2000. Aust. Inst. Mar. Sci., Townsville; 200 pp.