3532
IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 58, NO. 12, DECEMBER 2011
A Multiscale Framework Based on the Physiome Markup Languages for Exploring the Initiation of Osteoarthritis at the Bone–Cartilage Interface Vickie B. Shim*, Peter J. Hunter, Peter Pivonka, and Justin W. Fernandez
Abstract—The initiation of osteoarthritis (OA) has been linked to the onset and progression of pathologic mechanisms at the cartilage–bone interface. Most importantly, this degenerative disease involves cross-talk between the cartilage and subchondral bone environments, so an informative model should contain the complete complex. In order to evaluate this process, we have developed a multiscale model using the open-source ontologies developed for the Physiome Project with cartilage and bone descriptions at the cellular, micro, and macro levels. In this way, we can effectively model the influence of whole body loadings at the macro level and the influence of bone organization and architecture at the micro level, and have cell level processes that determine bone and cartilage remodeling. Cell information is then passed up the spatial scales to modify micro architecture and provide a macro spatial characterization of cartilage inflammation. We evaluate the framework by linking a common knee injury (anterior cruciate ligament deficiency) to proinflammatory mediators as a possible pathway to initiate OA. This framework provides a “virtual bone–cartilage” tool for evaluating hypotheses, treatment effects, and disease onset to inform and strengthen clinical studies. Index Terms—Cartilage, finite elements (FEs), multiscale, osteoarthritis (OA).
I. INTRODUCTION STEOARTHRITIS (OA) is a debilitating disease that causes widespread physical morbidity and impaired quality of life. Traditionally, OA has been regarded as the wear and tear of cartilage. However, recent evidence suggests that 1) OA is not just a disrupted cartilage mechanism but a breakdown of the whole cartilage–bone complex; hence, bone and cartilage should be treated together due to their intrinsic interrelationship; and
O
Manuscript received April 2, 2011; revised June 14, 2011; accepted August 1, 2011. Date of publication August 30, 2011; date of current version November 18, 2011. The work of V. Shim was supported by the Auckland University Faculty Research Development Funds and the Wishbone Trust Fund. The work of J. W. Fernandez was supported by the Auckland University Faculty Research Development Funds and the Aotearoa ABI Postdoctoral Fellowship. Asterisk indicates corresponding author. *V. Shim is with Auckland Bioengineering Institute, Auckland 1010, New Zealand (e-mail:
[email protected]). P. Hunter is with Auckland Bioengineering Institute, Auckland 1010, New Zealand (e-mail:
[email protected]). P. Pivonka is with School of Computer Science and Software Engineering, The University of Western Australia, Perth, W.A. 6009, Australia (e-mail: peter.
[email protected]). J. Fernandez is with Auckland Bioengineering Institute, Auckland 1010, New Zealand (e-mail:
[email protected]). Color versions of one or more of the figures in this paper are available online at http://ieeexplore.ieee.org. Digital Object Identifier 10.1109/TBME.2011.2165955
2) OA exists in the highly metabolic and inflammatory environments of adipose tissue, which control joint degradation or local inflammatory processes by secreting various proinflammatory cytokines [1]. Traditional biomechanical studies have focused on macro-level factors mainly, such as cartilage thinning or material property change. The question of whether degeneration in cartilage is initiated by changes in articular cartilage or the underlying subchondral bone is difficult to answer when one considers them in isolation. Therefore, our modeling approach aims to have a holistic view of bone–cartilage physiology with information viewed collectively rather than a fragmented approach. This study employs a multiscale framework developed as part of the Physiome Project and previously demonstrated for the heart [2]. In order to integrate these modeling scales, an efficient set of numerical tools is necessary to link, store models, and employ different methodologies at different scales. Here, we illustrate this structure for an important musculoskeletal problem, OA induced at the cartilage–bone interface. Moreover, numerous models of bone remodeling have been reported across different length scales and the need to link these using an open-source markup ontology has been recently suggested [3]. It is well accepted that bone remodeling is initiated by osteocyte apoptosis (cell death) either biomechanically or biochemically [4]. The human body is comprised of tissues that have formed to endure mechanical loads throughout life. Examining cartilage and bone, we can observe that there is transition zone of calcified cartilage between cartilage and bone, which provides a gradual mechanical transition of load from cartilage to bone. The aim of this study is to present a multiscale finite element (FE) framework of the cartilage bone junction in order to analyze the initiation of OA. This is accomplished by numerically linking information across spatial scales from a whole continuum knee model down to a model of cell mechanics described using ordinary differential equations. To initiate this process, we adopt a well-known condition, anterior cruciate ligament deficiency (ACLD), that leads to abnormal mechanical loading at the knee as a possible precursor [5]. Previous models have used FE analysis to show that both cartilage thinning and subchondral stiffening do not lead to significant changes in cartilage strain [6]. These studies suggest that altered mechanical loading is a likely candidate where normally unloaded cartilage now supports higher loads. A recent review also suggested that geometrical features such as bony spurs may have an influence but the effects are currently unknown [6]. To address this, we have developed a detailed micro model of the
0018-9294/$26.00 © 2011 IEEE
IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 58, NO. 12, DECEMBER 2011
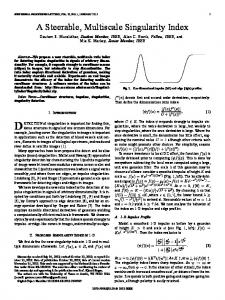
Fig. 1. Multiscale framework used in this study depicting the coupled links from the whole continuum knee model to the macro and cell level descriptions.
bone cartilage union that captures the subchondral architecture and the irregular shape at the bone–cartilage interface. We hypothesize that when the mechanical environment of the knee changes abnormally, such as with ACLD gait, bone remodeling will modify micro architecture and ultimately influence peak cartilage strains. These strains will then induce proinflammatory cytokines to degrade cartilage quality at the whole organ level. II. METHOD Fig. 1 outlines the multiscale approach developed for this study. An anatomically based knee model developed to investigate joint loading in gait was the starting point [8]. This model consisted of the femur, tibia, ligaments, menisci, and cartilage. The geometries were developed using high-order cubic Hermite elements, fitted to the Visible Human dataset and customized to a subject using free-form deformation methods [9]. The model assembly is detailed in AnatML [10], which describes the anatomical linkages for the whole human body and acts as a store for the anatomical geometries. For both the macro and micro models, the bones and cartilage were treated as deformable linear elastic materials [8], using a “St-Venant Kirchoff” constitutive law [11]. Macro and micro cartilage models had fiber information but it was not employed for this study (an isotropic assumption was used instead). The model components are described using FieldML [12], which captures in a generic web-based language the spatially vary-
3533
Fig. 2. (a) Electron microscope image showing cartilage, ZCC, and subchondral bone; (b) 2-D slice of undeformed FE mesh and (c) compressed FE mesh with 100 μm loading; (d) healthy von Mises strain and bone architecture; (e) ACLD von Mises strain and remodeled architecture after 3 months of bone evolution. Dark blue is 0 με, light blue is 1500 με, yellow is 3000 με, and red is 4500 με or greater; cartilage von Mises strain pattern at interface for (f) healthy and (g) ACLD remodeled bone. Dark blue is 4500 με, light blue is 6000 με, yellow is 7500 με, and red is 9000 με or greater
ing information for each model component. This includes the geometry, fiber information, material properties, and resulting stress/strain distributions after simulation. The micro FE model [see Fig. 2(a)] was adapted from the imaging study by Zizak et al. [13] with a depth of 450 μm for cartilage, 200 μm for the zone of calcified cartilage (ZCC), and 350 μm for subchondral bone. Young’s modulus and Poisson’s ratios were (E = 10 MPa, ν = 0.46) for cartilage, (E = 100 MPa, ν = 0.3) for ZCC, and (E = 1 GPa, ν = 0.3) for bone. To model the loads experienced by the knee we simulated gait, the most common human task. Two key loading points of the gait cycle were identified, heel-strike (HS) and contralateral toe-off (CTO), as representative of gait to load the macro model. Inverse dynamics was performed to obtain joint torques and forces from these two points. HS was modeled first followed by CTO and passed to the micro model. To simulate remodeling, we designated the healthy strain state (normal gait) as homeostatic equilibrium, where bone and cartilage are maintained and no remodeling occurs. We then simulated the altered cartilage strains due to loading from ACLD. The difference between the homeostatic and ACLD strains was the excitation for bone remodeling to initiate. Micro models of cartilage (seen as red cubes in Fig. 1) are spatially placed across the bone–cartilage interface to capture an average strain. The average strain in each cube is then passed to a micro model. The linking between the macro and micro models was integrated within our FE software CMISS (www.cmiss.org). This package is open source and facilitates the numerical linkages between the modeling scales.
3534
IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 58, NO. 12, DECEMBER 2011
For each micro model, the average strain was then applied to the surface of the detailed micro bone–cartilage model and we modeled the propagation through the cartilage, ZCC, and subchondral bone regions. The micro model was repeatedly run with each simulation representing a weeks’ worth of loading. After each simulation, the geometry was remodeled (through calling a cell level model) and the simulation run again with the new bone architecture producing new strain patterns in both the bone, ZCC and cartilage. Fig. 2 shows the remodeled bone geometry and associated strains after 3 months (under the assumption that it is only from walking). Our modeling framework accounts for the homogenization of material properties (at bone and cartilage) by passing the deformation gradient to the micro model and returning the average stress back to the whole organ but this was not the focus of this study. Rather, we were interested in whether bone remodeling induces cartilage strain change and excitation of a proinflammatory response. Hence, we did not rerun the macro model iteratively with new material properties and present them in this paper. The micro model was informed by two cell models. A bone remodeling algorithm [14] based on the RANK–RANKL–OPG pathway predicts the number of active osteoblasts (to deposit bone) and osteoclasts (to absorb bone) coded in CellML [15]. The cell model used micro strains as excitation to predict temporal concentrations of osteoblasts and osteoclasts over their life cycle, from precursor cells to active working cells and finally apoptosis. The osteocyte sensors are strategically placed at numerical integration points within each element and inform the FE nodes of the intensity of the strain signal based on their distance to the nodes. Each node then calls the CellML bone model to determine the osteoblast/osteoclast numbers to provide the rate of remodeling, which is directioned normal to each surface. Similarly, a cartilage damage prediction model was developed based on the work of Nam et al. [16], which gives a quantitative description of the action of nuclear factor-kappa B (NF-κB) signaling cascade under mechanical stimulation. Peak cartilage strains were used as excitation of the protein complex IκB kinase (IKK) which activates the NF-κB pathway, leading to induction of a number of proinflammatory genes. The simulation was run on our High Performance Computing facility (IBM X3850 with Intel Xeon X7560 2.66 GHz (24 cores) and 128 GB memory), taking 8 h to run the macro model and 20 min per micro model iteration. The cell and macro-level knee models have been uploaded to our web sites (www.cmiss.org and www.cellml.org) and are freely available for academic use. For this study, we examined the remodeled micro bone, altered strain pattern, and spatial mapping of inflammatory cartilage response at the macro level under ACLD loading and when normal loading is returned.
Fig. 3. Inflammatory response predicted in terms of the expression level of nitric oxide synthase (NOS2) for (a) ACLD and (b) under normal loading.
knee model, the tibiofemoral stress was shifted anteriorly and consistent with previous ACLD knee findings [18]. Fig. 2(a)–(c) illustrates a slice of the 3-D micro FE model and an illustrative micro cartilage deformation of 100 μm. Fig. 2(d) shows the healthy subchondral bone architecture and von Mises strain pattern considered to be in homeostatic equilibrium. After 3 months of simulated ACLD gait [see Fig. 2(e)], the abnormal loading evolves the bone pores in response to the pattern of strain excitation. In particular, the pores have reduced and the bone has thickened. The von Mises strain was used as this is a scalar accounting for all the principal components and a commonly used yield criteria, previously used for cortical bone remodeling [19]. It is clear that the subchondral bone architecture creates regions of strain shielding leading to a spatially varying strain excitation signal. This remodeling approach accounted for both bone growth and bone shape evolution enhancing the ZCC undulating profile and tidemark irregularity. This pattern resembles the finger-like structures that are reported to form during the progression of OA [7]. The peak strain is also 15–20% higher in the ACLD knee at the ZCC interface with higher strain closer to the ZCC interface [see Fig. 2(g)] compared to the normal knee [see Fig. 2(f)]. The increase in cartilage strains increased the production of inflammatory cytokines as shown in Fig. 3. Fig. 3 shows the pattern of proinflammatory initiation across the cartilage in the remodeled knee for the ACLD loading [see Fig. 3(a)] and normal loading [see Fig. 3(b)]. In the anterior cartilage, primarily in the medial compartment, the model predicted an inflammatory response highlighting sites of likely degeneration. What is most interesting from our model prediction is that even after a return to normal loading [see Fig. 3(b)] inflammation continued in the remodeled location. The remaining cartilage has undergone insignificant remodeling and can easily bear the normal load again with no inflammatory response. This indicates that once the pathologic state is initiated, the degenerative cycle may not be reversible. IV. DISCUSSION
III. RESULTS The previously developed whole knee model [8] was consistent with cartilage deformations and kinematics reported by a previous macro-level study [17]. In the ACLD variant of our
We have presented a numerical framework showing the linkages between AnatML, FieldML, and CellML. This has been used to evaluate a possible mechanism for the initiation of OA by considering the coupling of bone and cartilage. This was
IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 58, NO. 12, DECEMBER 2011
established by considering the excitation for bone remodeling being influenced by the complex subchondral bone architecture to induce a spatially varying remodeling pattern in the cartilage bone tidemark. This, in turn, leads to changes in peak cartilage strains that can induce proinflammatory responses. This is consistent with previously reported work that showed altered mechanical loads can lead to loading in more fibrillated and thinner regions of cartilage, which may initiate cartilage breakdown [5], [20]. The presented modeling framework is a web-based opensource repository of models, which contributing scientists can easily use and disseminate their findings. Furthermore, this framework can employ different numerical methods at different scales. For example, while the macro and micro models both employed FEs in this study, the micro model could easily employ a meshless method like smooth particle hydrodynamics [19], which can easily handle complicated evolving geometries during remodeling. The findings in this study may explain why altered or traumatic subchondral bone loadings can initiate a degenerative cycle in the cartilage matrix. Furthermore, the subchondral bone expansion and detection of complicated bony spurs are also consistent early signs of OA [21]. When altered loads were applied, the subchondral plate adapted. Therefore, loading due to misalignment plays a role in subchondral bone changes. Hence, thinning of the cartilage and stiffening of the subchondral bone may not be as significant as the location of the load [6]. These results should be considered with the fact that assuming highly accurate fluoroscopic data at the macro level (errors of 0.1–1.0 mm), input strains to the micro model can be up to 10% of the macro scale error. Sensitivity analysis showed a linear trend in the micro model when perturbed within the operating range of 0.05–0.2 mm with a 10% variation in load producing a 10% change in peak strain. The current implementation of this framework is the first step at linking key scales of bone and cartilage modeling. It primarily examines a biomechanical link between cartilage and bone; however, molecular cross-talk has been postulated [7]. It has been shown that vascular channels and the irregular geometry at the interface may facilitate molecular transport. One improvement to the model that may facilitate cross-talk is to link feedback between the bone and cartilage cell models. It has been suggested that the irregular geometry provides a mechanism of molecular diffusion from the bone through the ZCC to cartilage [7]. Currently, the model was only evaluated by comparing the predicted kinematics and joint contact pressure with previous macro studies [17]. We also found that micro-mechanical strain increase supported cartilage thinning, ZCC thickening, and the filamentary structures observed in imaging studies of pathologic cartilage [21]. This study also used the same micro model at all the spatial locations of the bone. Ideally, unique micro bone models will generate spatially varying remodeling responses. Second, the current study is only run with loading from gait; however, including more strenuous activities such as stair-climbing and squatting can provide a more representative homeostatic condition for normal loading. Third, the inputs ap-
3535
plied to the knee model did not reflect the compensatory gait mechanics and muscle activation patterns observed in many ACLD patients. That is, the joint angles, ground forces, muscle forces, and joint-reaction forces input to the ACLD model were assumed to be identical with those present during normal gait. In conclusion, we have demonstrated the first attempt at linking three scales together to explain a possible mechanism for the initiation of OA. The use of the Physiome Project infrastructure has made linking the scales more efficient, and the markup languages (AnatML, FieldML, and CellML) provide a structure for easy model storage and dissemination. REFERENCES [1] R. M. Aspden, B. A. Scheven, and J. D. Hutchison, “Osteoarthritis as a systemic disorder including stromal cell differentiation and lipid metabolism,” Lancet, vol. 357, pp. 1118–1120, Apr. 2001. [2] P. J. Hunter and T. K. Borg, “Integration from proteins to organs: The Physiome Project,” Nat. Rev. Mol. Cell Biol., vol. 4, pp. 237–243, Mar. 2003. [3] F. A. Gerhard, D. J. Webster, G. H. van Lenthe, and R. Muller, “In silico biology of bone modelling and remodelling: Adaptation,” Philos. Trans. A Math. Phys. Eng. Sci., vol. 367, pp. 2011–2030, May 2009. [4] T. J. Martin and E. Seeman, “Bone remodelling: Its local regulation and the emergence of bone fragility,” Best Pract. Res. Clin. Endocrinol. Metab., vol. 22, pp. 701–722, Oct. 2008. [5] T. P. Andriacchi, S. Koo, and S. F. Scanlan, “Gait mechanics influence healthy cartilage morphology and osteoarthritis of the knee,” J. Bone Joint Surgery, vol. 91A, pp. 95–101, Feb. 2009. [6] P. R. van den Broek, An explanation for changes in subchondral bone during osteoarthritis,” Master’s thesis, Dept. of Biomechan. Eng., Eindhoven Univ. of Technology, Eindhoven, the Netherlands, 2006. [7] R. J. Lories and F. P. Luyten, “The bone-cartilage unit in osteoarthritis,” Nat. Rev. Rheumatol., vol. 7, pp. 43–49, Jan. 2011. [8] V. B. Shim, K. Mithraratne, I. A. Anderson, and P. J. Hunter, “Simulating in vivo knee kinetics and kinematics of tibio-femoral articulation with a subject-specific finite element model,” in World Congress on Medical Physics and Biomedical Engineering, Sept. 7–12, 2009, O. D¨ossel and W. C. Schlegel, Eds., Munich, Germany: Springer Berlin Heidelberg, 2010, vol. 25/4, pp. 2315–2318. [9] J. W. Fernandez, P. Mithraratne, S. F. Thrupp, M. H. Tawhai, and P. J. Hunter, “Anatomically based geometric modelling of the musculoskeletal system and other organs,” Biomech. Model Mechanobiol., vol. 2, pp. 139–155, Mar. 2004. [10] P. Hunter, N. Smith, J. Fernandez, and M. Tawhai, “Integration from proteins to organs: the IUPS Physiome Project,” Mech. Ageing Dev., vol. 126, pp. 187–192, Jan. 2005. [11] J. W. Fernandez and P. J. Hunter, “An anatomically based patient-specific finite element model of patella articulation: towards a diagnostic tool,” Biomech. Model Mechanobiol., vol. 4, pp. 20–38, Aug. 2005. [12] G. R. Christie, P. M. Nielsen, S. A. Blackett, C. P. Bradley, and P. J. Hunter, “FieldML: Concepts and implementation,” Philos. Trans. A Math. Phys. Eng. Sci., vol. 367, pp. 1869–1884, May 28, 2009. [13] I. Zizak, P. Roschger, O. Paris, B. M. Misof, A. Berzlanovich, S. Bernstorff, H. Amenitsch, K. Klaushofer, and P. Fratzl, “Characteristics of mineral particles in the human bone/cartilage interface,” J. Struct. Biol., vol. 141, no. 3, pp. 208–217, Mar. 2003. [14] P. Pivonka, J. Zimak, D. W. Smith, B. S. Gardiner, C. R. Dunstan, N. A. Sims, T. J. Martin, and G. R. Mundy, “Model structure and control of bone remodeling: A theoretical study,” Bone, vol. 43, no. 2, pp. 249–263, Aug. 2008. [15] D. Nickerson and P. Hunter, “Using CellML in computational models of multiscale physiology,” in Proc. IEEE Int. Conf. Eng. Med. Biol. Soc., 2005, vol. 6, pp. 6096–6099. [16] J. Nam, B. D. Aguda, B. Rath, and S. Agarwal, “Biomechanical thresholds regulate inflammation through the NF-kappaB pathway: Experiments and modeling,” PLoS One, vol. 4, e5262, 2009. [17] F. Liu, M. Kozanek, A. Hosseini, S. K. Van de Velde, T. J. Gill, H. E. Rubash, and G. Li, “In vivo tibiofemoral cartilage deformation during the stance phase of gait,” J. Biomech., vol. 43, no. 4, pp. 658–665, Mar. 2010.
3536
IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 58, NO. 12, DECEMBER 2011
[18] T. P. Andriacchi, P. L. Briant, S. L. Bevill, and S. Koo, “Rotational changes at the knee after ACL injury cause cartilage thinning,” Clin. Orthopaedics Relat. Res., pp. 39–44, Jan. 2006. [19] J. Fernandez, R. Das, D. Thomas, P. Cleary, M. Sinnott, and J. Clement, “Strain reduction between cortical pore structures leads to bone weakening and fracture susceptibility: An investigation using smooth particle hydrodynamics,” in Proc. 6th World Congr. Biomechanics, Singapore, 2010, pp. 784–787.
[20] J. W. Fernandez, M. Akbarshahi, K. M. Crossley, K. B. Shelburne, and M. G. Pandy, “Model predictions of increased knee joint loading in regions of thinner articular cartilage after patellar tendon adhesion,” J. Orthop. Res., vol. 29, pp. 1168–1177, Mar. 2011. [21] A. Thambyah and N. Broom, “On how degeneration influences loadbearing in the cartilage-bone system: A microstructural and micromechanical study,” Osteoarthritis Cartilage, vol. 15, pp. 1410–1423, Dec. 2007.